

Antisense transcription in gammaretroviruses as a mechanism of insertional activation of host genes
- PMID: 20130045
- PMCID: PMC2849499
- DOI: 10.1128/JVI.02088-09
Antisense transcription in gammaretroviruses as a mechanism of insertional activation of host genes
Abstract
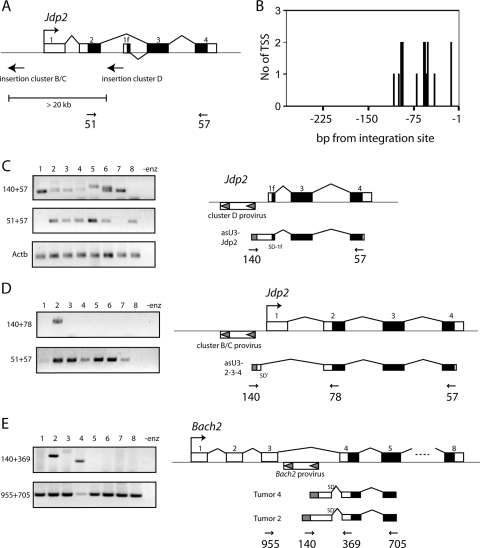
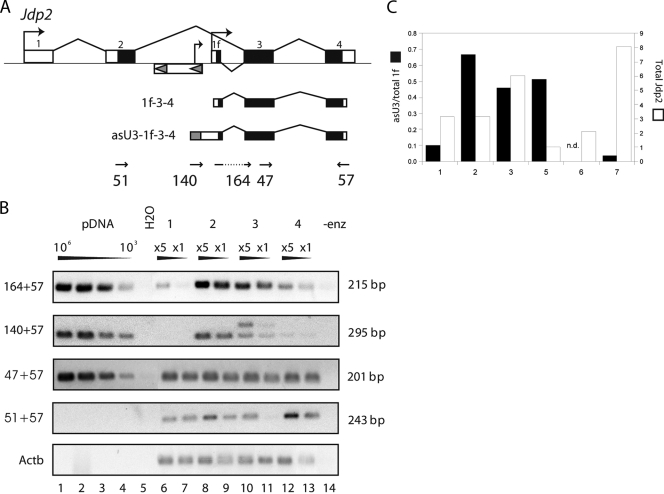
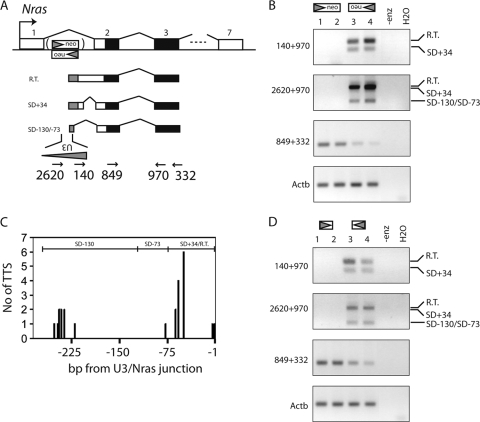
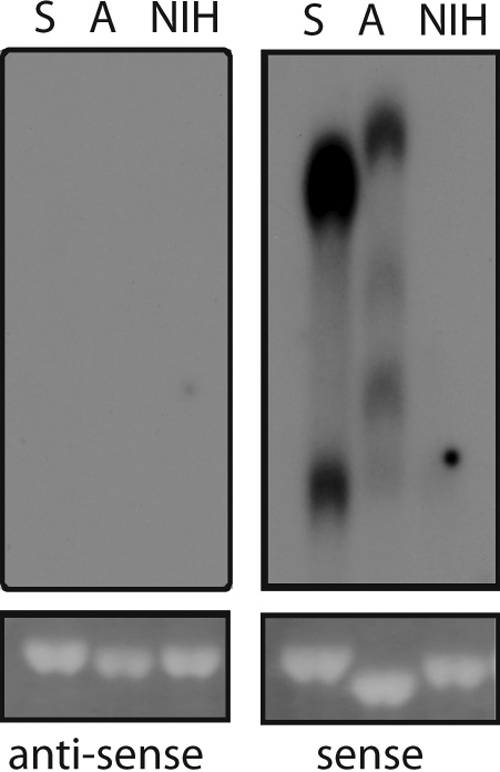
Transcription of retroviruses is initiated at the U3-R region boundary in the integrated provirus and continues unidirectionally to produce genomic and mRNA products of positive polarity. Several studies have recently demonstrated the existence of naturally occurring protein-encoding transcripts of negative polarity in complex retroviruses. We report here on the identification of transcripts of negative polarity in simple murine leukemia virus (MLV). In T-cell and B-cell lymphomas induced by SL3-3 and Akv MLV, antisense transcripts initiated in the U3 region of the proviral 5' long terminal repeat (LTR) and continued into the cellular proto-oncogenes Jdp2 and Bach2 to create chimeric transcripts consisting of viral and host sequence. The phenomenon was validated in vivo using a knock-in mouse model homozygous for a single LTR at a position known to activate Nras in B-cell lymphomas. A 5' rapid amplification of cDNA ends (RACE) analysis indicated a broad spectrum of initiation sites within the U3 region of the 5' LTR. Our data show for the first time transcriptional activity of negative polarity initiating in the U3 region of simple retroviruses and suggest a novel mechanism of insertional activation of host genes. Elucidation of the nature and potential regulatory role of 5' LTR antisense transcription will be relevant to the design of therapeutic vectors and may contribute to the increasing recognition of pervasive eukaryotic transcription.
Figures
Similar articles
-
Tumor model-specific proviral insertional mutagenesis of the Fos/Jdp2/Batf locus.Virology. 2005 Jul 5;337(2):353-64. doi: 10.1016/j.virol.2005.04.027. Virology. 2005. PMID: 15913695
-
Novel principles of gamma-retroviral insertional transcription activation in murine leukemia virus-induced end-stage tumors.Retrovirology. 2014 May 19;11:36. doi: 10.1186/1742-4690-11-36. Retrovirology. 2014. PMID: 24886479 Free PMC article.
-
Capture of a cellular transcriptional unit by a retrovirus: mode of provirus activation in embryonal carcinoma cells.J Virol. 1992 Aug;66(8):4982-91. doi: 10.1128/JVI.66.8.4982-4991.1992. J Virol. 1992. PMID: 1321282 Free PMC article.
-
Tag, you're hit: retroviral insertions identify genes involved in cancer.Trends Mol Med. 2003 Feb;9(2):43-5. doi: 10.1016/s1471-4914(03)00003-0. Trends Mol Med. 2003. PMID: 12615036 Review.
-
Virus-host interactions and the pathogenesis of murine and human oncogenic retroviruses.Curr Top Microbiol Immunol. 1991;171:95-171. doi: 10.1007/978-3-642-76524-7_5. Curr Top Microbiol Immunol. 1991. PMID: 1667631 Review. No abstract available.
Cited by
-
Human endogenous retroviruses sustain complex and cooperative regulation of gene-containing loci and unannotated megabase-sized regions.Retrovirology. 2015 Apr 17;12:32. doi: 10.1186/s12977-015-0161-9. Retrovirology. 2015. PMID: 25927889 Free PMC article.
-
Retroviral Antisense Transcripts and Genes: 33 Years after First Predicted, a Silent Retroviral Revolution?Viruses. 2021 Nov 4;13(11):2221. doi: 10.3390/v13112221. Viruses. 2021. PMID: 34835027 Free PMC article. Review.
-
Permissive Sense and Antisense Transcription from the 5' and 3' Long Terminal Repeats of Human T-Cell Leukemia Virus Type 1.J Virol. 2016 Jan 20;90(7):3600-10. doi: 10.1128/JVI.02634-15. J Virol. 2016. PMID: 26792732 Free PMC article.
-
Polarized expression of the membrane ASP protein derived from HIV-1 antisense transcription in T cells.Retrovirology. 2011 Sep 19;8:74. doi: 10.1186/1742-4690-8-74. Retrovirology. 2011. PMID: 21929758 Free PMC article.
-
Deregulated Nras expression in knock-in animals harboring a gammaretroviral long terminal repeat at the Nras/Csde1 locus.PLoS One. 2013;8(2):e56029. doi: 10.1371/journal.pone.0056029. Epub 2013 Feb 13. PLoS One. 2013. PMID: 23418499 Free PMC article.
References
-
- Birney, E., J. A. Stamatoyannopoulos, A. Dutta, R. Guigo, T. R. Gingeras, E. H. Margulies, Z. Weng, M. Snyder, E. T. Dermitzakis, R. E. Thurman, M. S. Kuehn, C. M. Taylor, S. Neph, C. M. Koch, S. Asthana, A. Malhotra, I. Adzhubei, J. A. Greenbaum, R. M. Andrews, P. Flicek, P. J. Boyle, H. Cao, N. P. Carter, G. K. Clelland, S. Davis, N. Day, P. Dhami, S. C. Dillon, M. O. Dorschner, H. Fiegler, P. G. Giresi, J. Goldy, M. Hawrylycz, A. Haydock, R. Humbert, K. D. James, B. E. Johnson, E. M. Johnson, T. T. Frum, E. R. Rosenzweig, N. Karnani, K. Lee, G. C. Lefebvre, P. A. Navas, F. Neri, S. C. Parker, P. J. Sabo, R. Sandstrom, A. Shafer, D. Vetrie, M. Weaver, S. Wilcox, M. Yu, F. S. Collins, J. Dekker, J. D. Lieb, T. D. Tullius, G. E. Crawford, S. Sunyaev, W. S. Noble, I. Dunham, F. Denoeud, A. Reymond, P. Kapranov, J. Rozowsky, D. Zheng, R. Castelo, A. Frankish, J. Harrow, S. Ghosh, A. Sandelin, I. L. Hofacker, R. Baertsch, D. Keefe, S. Dike, J. Cheng, H. A. Hirsch, E. A. Sekinger, J. Lagarde, J. F. Abril, A. Shahab, C. Flamm, C. Fried, J. Hackermuller, J. Hertel, M. Lindemeyer, K. Missal, A. Tanzer, S. Washietl, J. Korbel, O. Emanuelsson, J. S. Pedersen, N. Holroyd, R. Taylor, D. Swarbreck, N. Matthews, M. C. Dickson, D. J. Thomas, M. T. Weirauch, J. Gilbert, et al. 2007. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 447:799-816. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
