

Preparation and topology of the Mediator middle module
- PMID: 20123732
- PMCID: PMC2879511
- DOI: 10.1093/nar/gkq029
Preparation and topology of the Mediator middle module
Abstract
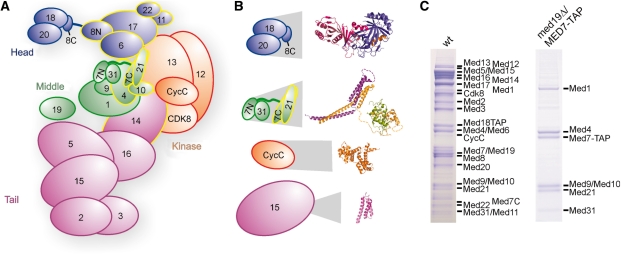
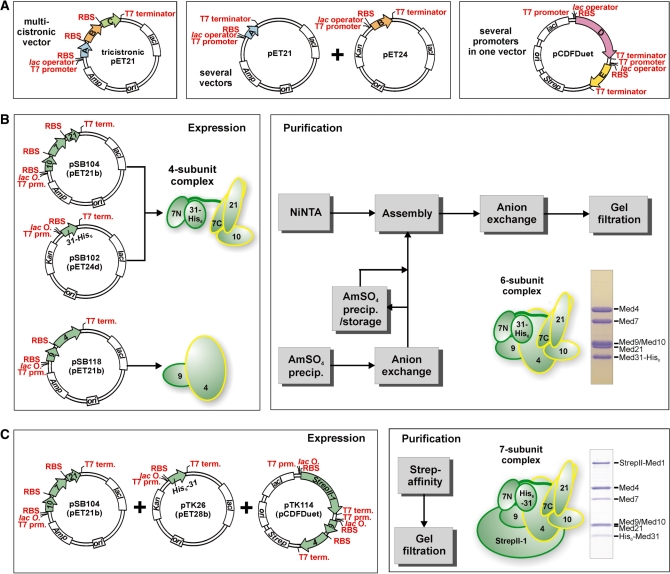
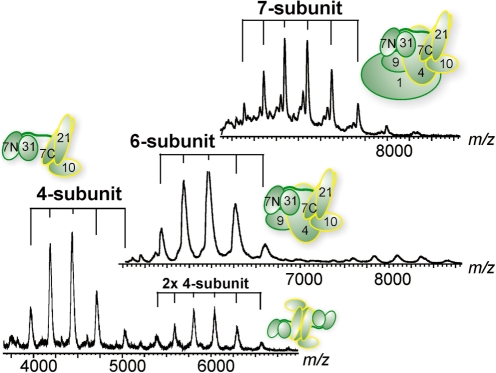
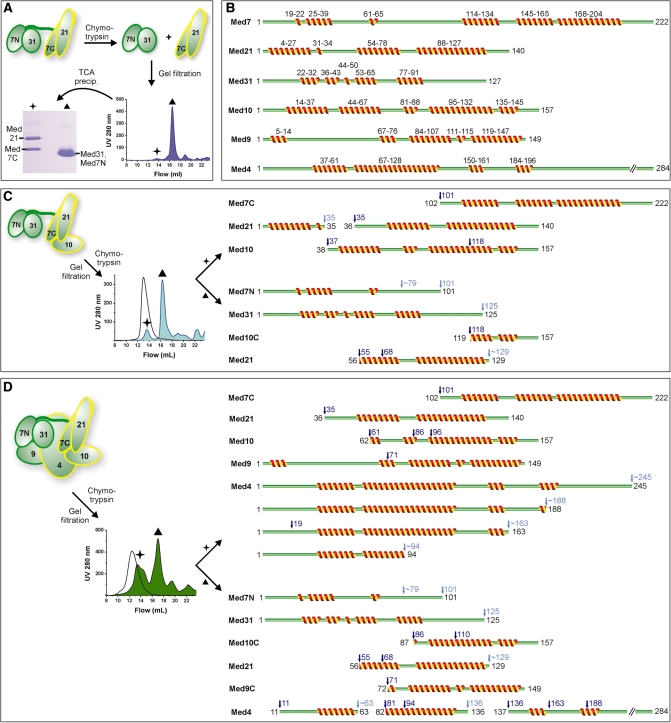
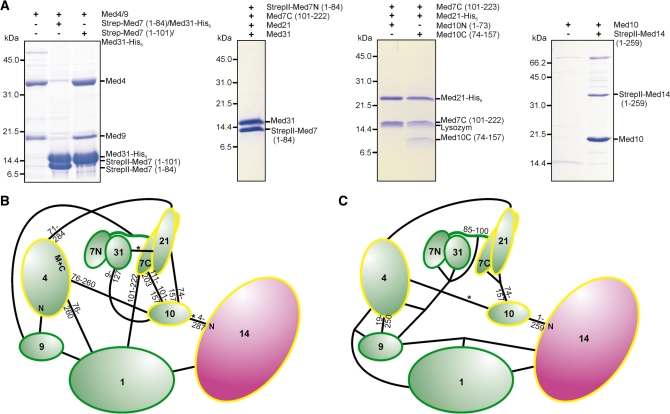
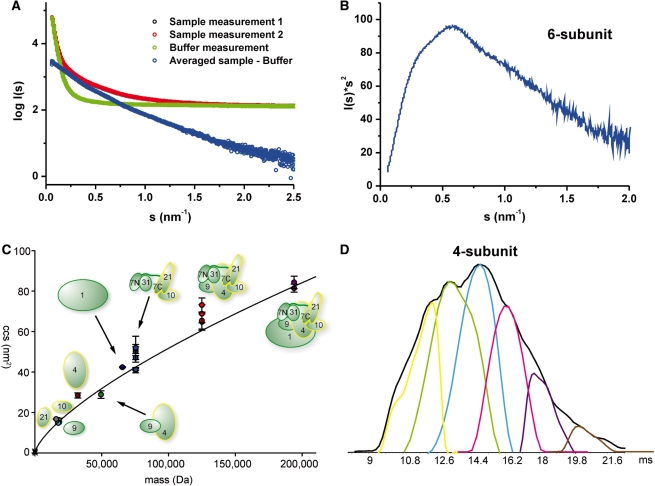
Mediator is the central coactivator complex required for regulated transcription by RNA polymerase (Pol) II. Mediator consists of 25 subunits arranged in the head, middle, tail and kinase modules. Structural and functional studies of Mediator are limited by the availability of protocols for the preparation of recombinant modules. Here, we describe protocols for obtaining pure endogenous and recombinant complete Mediator middle module from Saccharomyces cerevisiae that consists of seven subunits: Med1, 4, 7, 9, 10, 21 and 31. Native mass spectrometry reveals that all subunits are present in equimolar stoichiometry. Ion-mobility mass spectrometry, limited proteolysis, light scattering and small-angle X-ray scattering all indicate a high degree of intrinsic flexibility and an elongated shape of the middle module. Protein-protein interaction assays combined with previously published data suggest that the Med7 and Med4 subunits serve as a binding platform to form the three heterodimeric subcomplexes, Med7N/21, Med7C/31 and Med4/9. The subunits, Med1 and Med10, which bridge to the Mediator tail module, bind to both Med7 and Med4.
Figures
Similar articles
-
Model of the Mediator middle module based on protein cross-linking.Nucleic Acids Res. 2013 Nov;41(20):9266-73. doi: 10.1093/nar/gkt704. Epub 2013 Aug 11. Nucleic Acids Res. 2013. PMID: 23939621 Free PMC article.
-
Structure of the Mediator head module.Nature. 2012 Dec 20;492(7429):448-51. doi: 10.1038/nature11670. Epub 2012 Oct 31. Nature. 2012. PMID: 23123849
-
Facilitated assembly of the preinitiation complex by separated tail and head/middle modules of the mediator.J Mol Biol. 2012 Jan 20;415(3):464-74. doi: 10.1016/j.jmb.2011.11.020. Epub 2011 Nov 23. J Mol Biol. 2012. PMID: 22137896 Free PMC article.
-
Origins and activity of the Mediator complex.Semin Cell Dev Biol. 2011 Sep;22(7):729-34. doi: 10.1016/j.semcdb.2011.07.021. Epub 2011 Jul 28. Semin Cell Dev Biol. 2011. PMID: 21821140 Free PMC article. Review.
-
Twenty years of Mediator complex structural studies.Biochem Soc Trans. 2019 Feb 28;47(1):399-410. doi: 10.1042/BST20180608. Epub 2019 Feb 7. Biochem Soc Trans. 2019. PMID: 30733343 Free PMC article. Review.
Cited by
-
Subunit architecture and functional modular rearrangements of the transcriptional mediator complex.Cell. 2014 Jun 5;157(6):1430-1444. doi: 10.1016/j.cell.2014.05.015. Epub 2014 May 29. Cell. 2014. PMID: 24882805 Free PMC article.
-
Core Mediator structure at 3.4 Å extends model of transcription initiation complex.Nature. 2017 May 11;545(7653):248-251. doi: 10.1038/nature22328. Epub 2017 May 3. Nature. 2017. PMID: 28467824
-
The diverse and expanding role of mass spectrometry in structural and molecular biology.EMBO J. 2016 Dec 15;35(24):2634-2657. doi: 10.15252/embj.201694818. Epub 2016 Oct 26. EMBO J. 2016. PMID: 27797822 Free PMC article. Review.
-
Mass spectrometric analysis of intact human monoclonal antibody aggregates fractionated by size-exclusion chromatography.Pharm Res. 2010 Oct;27(10):2197-204. doi: 10.1007/s11095-010-0224-5. Epub 2010 Aug 3. Pharm Res. 2010. PMID: 20680668 Free PMC article.
-
Role of integrative structural biology in understanding transcriptional initiation.Methods. 2019 Apr 15;159-160:4-22. doi: 10.1016/j.ymeth.2019.03.009. Epub 2019 Mar 16. Methods. 2019. PMID: 30890443 Free PMC article. Review.
References
-
- Bjorklund S, Gustafsson CM. The yeast Mediator complex and its regulation. Trends Biochem Sci. 2005;30:240–244. - PubMed
-
- Kornberg RD. Mediator and the mechanism of transcriptional activation. Trends Biochem Sci. 2005;30:235–239. - PubMed
-
- Malik S, Roeder RG. Transcriptional regulation through Mediator-like coactivators in yeast and metazoan cells. Trends Biochem Sci. 2000;25:277–283. - PubMed
-
- Naar AM, Lemon BD, Tjian R. Transcriptional coactivator complexes. Annu. Rev. Biochem. 2001;70:475–501. - PubMed
-
- Asturias FJ, Jiang YW, Myers LC, Gustafsson CM, Kornberg RD. Conserved structures of mediator and RNA polymerase II holoenzyme. Science. 1999;283:985–987. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
