

Evidence for a novel coding sequence overlapping the 5'-terminal approximately 90 codons of the gill-associated and yellow head okavirus envelope glycoprotein gene
- PMID: 20017924
- PMCID: PMC2805633
- DOI: 10.1186/1743-422X-6-222
Evidence for a novel coding sequence overlapping the 5'-terminal approximately 90 codons of the gill-associated and yellow head okavirus envelope glycoprotein gene
Abstract
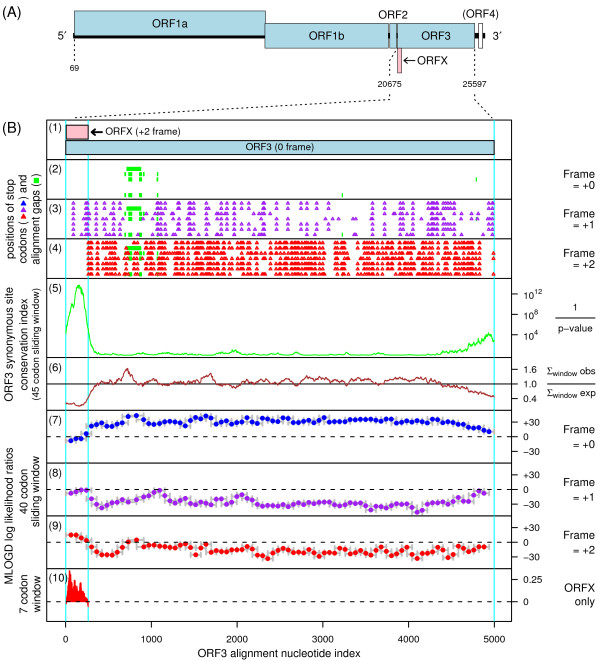
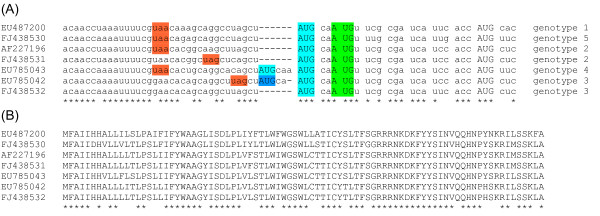
The genus Okavirus (order Nidovirales) includes a number of viruses that infect crustaceans, causing major losses in the shrimp industry. These viruses have a linear positive-sense ssRNA genome of approximately 26-27 kb, encoding a large replicase polyprotein that is expressed from the genomic RNA, and several additional proteins that are expressed from a nested set of 3'-coterminal subgenomic RNAs. In this brief report, we describe the bioinformatic discovery of a new, apparently coding, ORF that overlaps the 5' end of the envelope glycoprotein encoding sequence, ORF3, in the +2 reading frame. The new ORF has a strong coding signature and, in fact, is more conserved at the amino acid level than the overlapping region of ORF3. We propose that translation of the new ORF initiates at a conserved AUG codon separated by just 2 nt from the ORF3 AUG initiation codon, resulting in a novel 86 amino acid protein.
Figures
Similar articles
-
A case for a CUG-initiated coding sequence overlapping torovirus ORF1a and encoding a novel 30 kDa product.Virol J. 2009 Sep 8;6:136. doi: 10.1186/1743-422X-6-136. Virol J. 2009. PMID: 19737402 Free PMC article.
-
RNA transcription analysis and completion of the genome sequence of yellow head nidovirus.Virus Res. 2008 Sep;136(1-2):157-65. doi: 10.1016/j.virusres.2008.05.008. Epub 2008 Jun 11. Virus Res. 2008. PMID: 18582978 Free PMC article.
-
ICTV Virus Taxonomy Profile: Roniviridae.J Gen Virol. 2021 Jan;102(1):jgv001514. doi: 10.1099/jgv.0.001514. J Gen Virol. 2021. PMID: 33108263 Free PMC article. Review.
-
A virulent isolate of yellow head nidovirus contains a deformed envelope glycoprotein gp116.Virology. 2009 Feb 5;384(1):192-200. doi: 10.1016/j.virol.2008.10.042. Epub 2008 Dec 2. Virology. 2009. PMID: 19049843
-
Genomic organization, biology, and diagnosis of Taura syndrome virus and yellowhead virus of penaeid shrimp.Adv Virus Res. 2004;63:353-421. doi: 10.1016/S0065-3527(04)63006-5. Adv Virus Res. 2004. PMID: 15530565 Free PMC article. Review. No abstract available.
Cited by
-
Evolution of viral proteins originated de novo by overprinting.Mol Biol Evol. 2012 Dec;29(12):3767-80. doi: 10.1093/molbev/mss179. Epub 2012 Jul 19. Mol Biol Evol. 2012. PMID: 22821011 Free PMC article.
-
Horizontal gene transfers with or without cell fusions in all categories of the living matter.Adv Exp Med Biol. 2011;714:5-89. doi: 10.1007/978-94-007-0782-5_2. Adv Exp Med Biol. 2011. PMID: 21506007 Free PMC article. Review.
-
An alternative -1/+2 open reading frame exists within viral N(pro)(1-19) region of bovine viral diarrhea virus SD-1.Virus Res. 2012 Jan;163(1):341-51. doi: 10.1016/j.virusres.2011.10.022. Epub 2011 Nov 4. Virus Res. 2012. PMID: 22079882 Free PMC article.
-
Interspecific adaptation by binary choice at de novo polyomavirus T antigen site through accelerated codon-constrained Val-Ala toggling within an intrinsically disordered region.Nucleic Acids Res. 2015 May 26;43(10):4800-13. doi: 10.1093/nar/gkv378. Epub 2015 Apr 22. Nucleic Acids Res. 2015. PMID: 25904630 Free PMC article.
References
-
- Cowley JA, Dimmock CM, Walker PJ. Gill-associated nidovirus of Penaeus monodon prawns transcribes 3'-coterminal subgenomic mRNAs that do not possess 5'-leader sequences. J Gen Virol. 2002;83:927–935. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
