

Hierarchical rules for Argonaute loading in Drosophila
- PMID: 19917252
- PMCID: PMC2795325
- DOI: 10.1016/j.molcel.2009.09.028
Hierarchical rules for Argonaute loading in Drosophila
Abstract
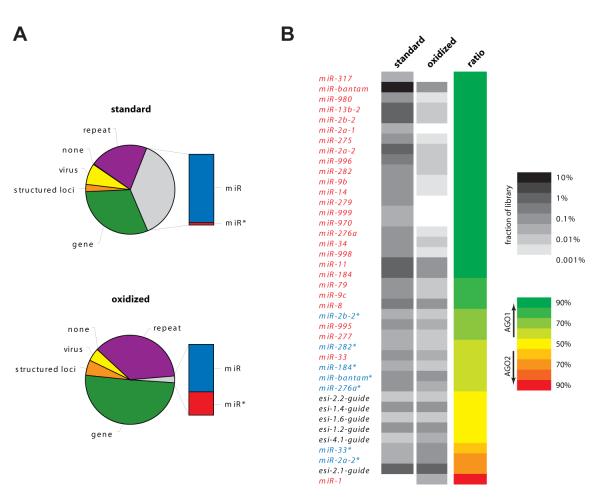
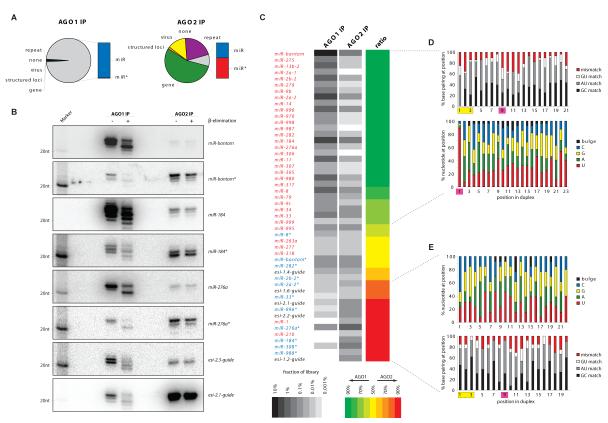
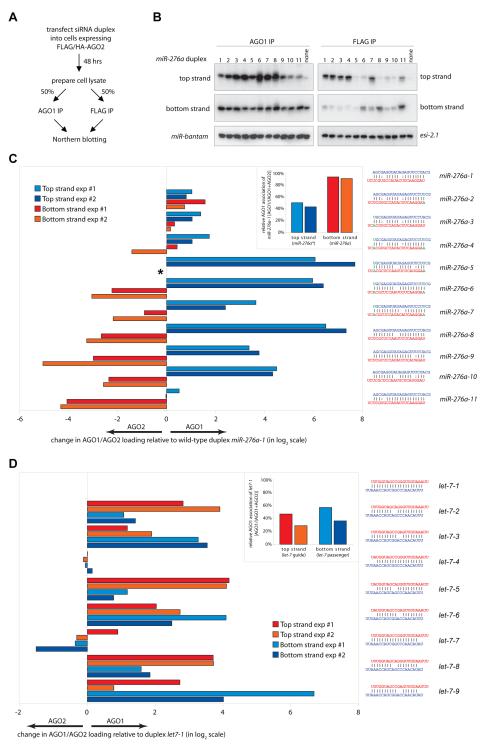
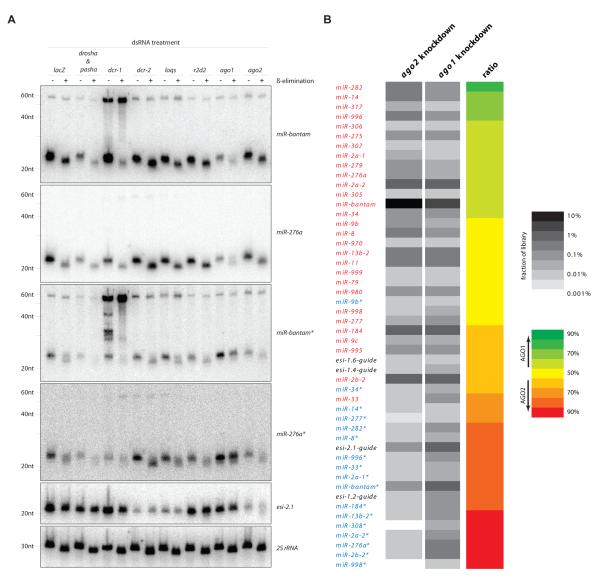
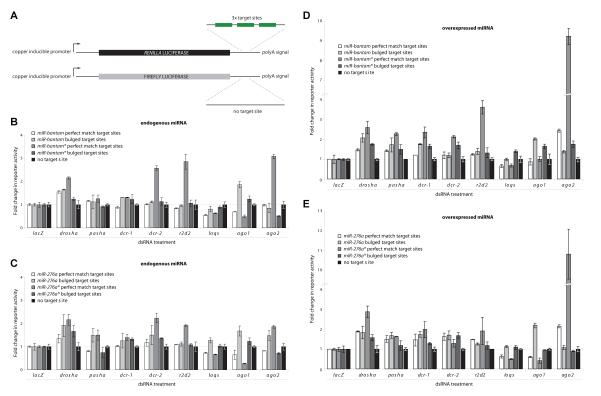
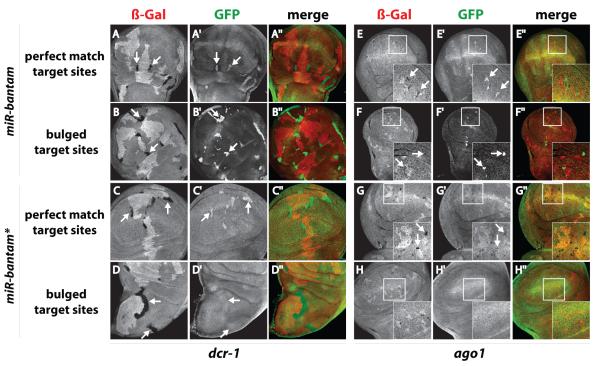
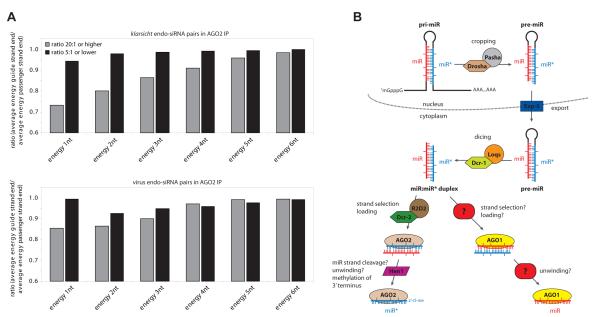
Drosophila Argonaute-1 and Argonaute-2 differ in function and small RNA content. AGO2 binds to siRNAs, whereas AGO1 is almost exclusively occupied by microRNAs. MicroRNA duplexes are intrinsically asymmetric, with one strand, the miR strand, preferentially entering AGO1 to recognize and regulate the expression of target mRNAs. The other strand, miR*, has been viewed as a byproduct of microRNA biogenesis. Here, we show that miR*s are often loaded as functional species into AGO2. This indicates that each microRNA precursor can potentially produce two mature small RNA strands that are differentially sorted within the RNAi pathway. miR* biogenesis depends upon the canonical microRNA pathway, but loading into AGO2 is mediated by factors traditionally dedicated to siRNAs. By inferring and validating hierarchical rules that predict differential AGO loading, we find that intrinsic determinants, including structural and thermodynamic properties of the processed duplex, regulate the fate of each RNA strand within the RNAi pathway.
Figures
Similar articles
-
Distinct mechanisms for microRNA strand selection by Drosophila Argonautes.Mol Cell. 2009 Nov 13;36(3):431-44. doi: 10.1016/j.molcel.2009.09.027. Mol Cell. 2009. PMID: 19917251 Free PMC article.
-
Sorting of Drosophila small silencing RNAs.Cell. 2007 Jul 27;130(2):299-308. doi: 10.1016/j.cell.2007.05.057. Cell. 2007. PMID: 17662944 Free PMC article.
-
Target RNA-directed tailing and trimming purifies the sorting of endo-siRNAs between the two Drosophila Argonaute proteins.RNA. 2011 Jan;17(1):54-63. doi: 10.1261/rna.2498411. Epub 2010 Nov 24. RNA. 2011. PMID: 21106652 Free PMC article.
-
Argonaute-mediated translational repression (and activation).Fly (Austin). 2009 Jul-Sep;3(3):204-6. Epub 2009 Jul 14. Fly (Austin). 2009. PMID: 19556851 Review.
-
The RNAi pathway initiated by Dicer-2 in Drosophila.Cold Spring Harb Symp Quant Biol. 2006;71:39-44. doi: 10.1101/sqb.2006.71.008. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381278 Review.
Cited by
-
Multiple sensors ensure guide strand selection in human RNAi pathways.RNA. 2013 May;19(5):639-48. doi: 10.1261/rna.037424.112. Epub 2013 Mar 26. RNA. 2013. PMID: 23531496 Free PMC article.
-
Probing the microRNA pathway with small molecules.Bioorg Med Chem. 2013 Oct 15;21(20):6119-23. doi: 10.1016/j.bmc.2013.05.030. Epub 2013 May 25. Bioorg Med Chem. 2013. PMID: 23791866 Free PMC article. Review.
-
Homeostatic control of Argonaute stability by microRNA availability.Nat Struct Mol Biol. 2013 Jul;20(7):789-95. doi: 10.1038/nsmb.2606. Epub 2013 May 26. Nat Struct Mol Biol. 2013. PMID: 23708604 Free PMC article.
-
Target RNA-directed trimming and tailing of small silencing RNAs.Science. 2010 Jun 18;328(5985):1534-9. doi: 10.1126/science.1187058. Science. 2010. PMID: 20558712 Free PMC article.
-
A 5'-uridine amplifies miRNA/miRNA* asymmetry in Drosophila by promoting RNA-induced silencing complex formation.Silence. 2011 Jun 7;2:4. doi: 10.1186/1758-907X-2-4. Silence. 2011. PMID: 21649885 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
