

piggyBac transposon/transposase system to generate CD19-specific T cells for the treatment of B-lineage malignancies
- PMID: 19905893
- PMCID: PMC2938363
- DOI: 10.1089/hum.2009.114
piggyBac transposon/transposase system to generate CD19-specific T cells for the treatment of B-lineage malignancies
Abstract
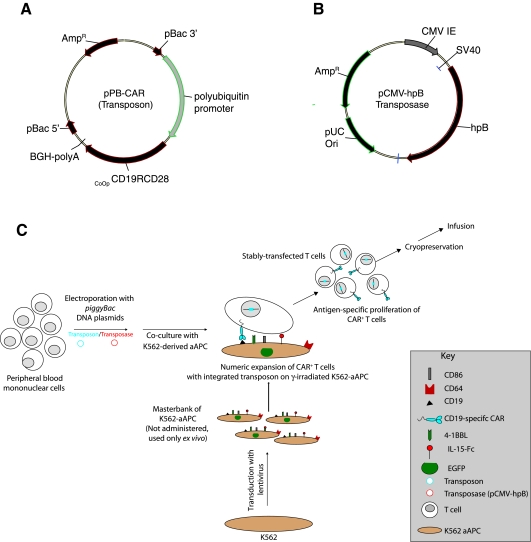
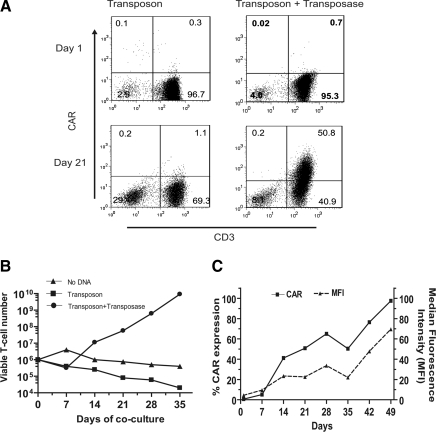
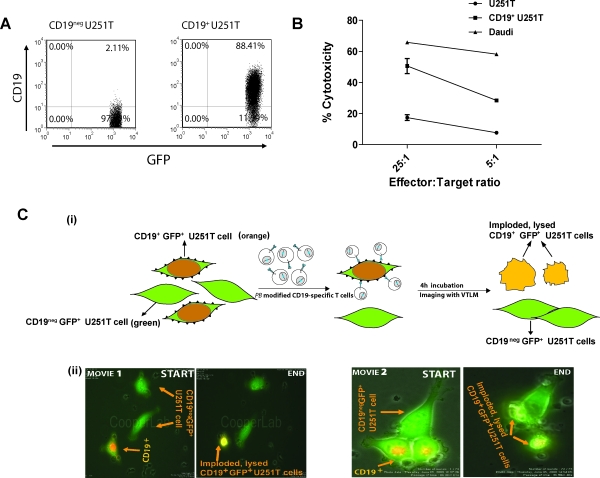
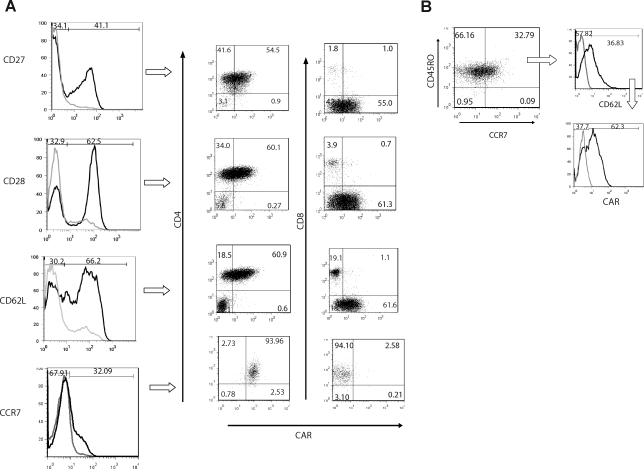
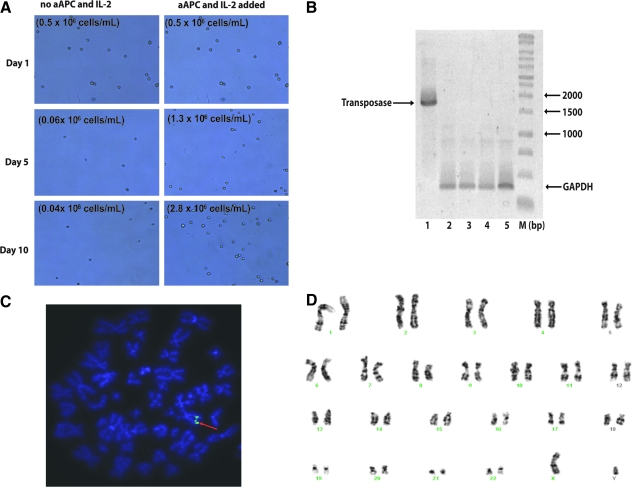
Nonviral integrating vectors can be used for expression of therapeutic genes. piggyBac (PB), a transposon/transposase system, has been used to efficiently generate induced pluripotent stems cells from somatic cells, without genetic alteration. In this paper, we apply PB transposition to express a chimeric antigen receptor (CAR) in primary human T cells. We demonstrate that T cells electroporated to introduce the PB transposon and transposase stably express CD19-specific CAR and when cultured on CD19(+) artificial antigen-presenting cells, numerically expand in a CAR-dependent manner, display a phenotype associated with both memory and effector T cell populations, and exhibit CD19-dependent killing of tumor targets. Integration of the PB transposon expressing CAR was not associated with genotoxicity, based on chromosome analysis. PB transposition for generating human T cells with redirected specificity to a desired target such as CD19 is a new genetic approach with therapeutic implications.
Figures
Similar articles
-
Manufacture of clinical-grade CD19-specific T cells stably expressing chimeric antigen receptor using Sleeping Beauty system and artificial antigen presenting cells.PLoS One. 2013 May 31;8(5):e64138. doi: 10.1371/journal.pone.0064138. Print 2013. PLoS One. 2013. PMID: 23741305 Free PMC article.
-
The hyperactive Sleeping Beauty transposase SB100X improves the genetic modification of T cells to express a chimeric antigen receptor.Gene Ther. 2011 Sep;18(9):849-56. doi: 10.1038/gt.2011.40. Epub 2011 Mar 31. Gene Ther. 2011. PMID: 21451576 Free PMC article.
-
The Tol2 transposon system mediates the genetic engineering of T-cells with CD19-specific chimeric antigen receptors for B-cell malignancies.Gene Ther. 2015 Feb;22(2):209-15. doi: 10.1038/gt.2014.104. Epub 2014 Nov 27. Gene Ther. 2015. PMID: 25427612 Free PMC article.
-
A transposon and transposase system for human application.Mol Ther. 2010 Apr;18(4):674-83. doi: 10.1038/mt.2010.2. Epub 2010 Jan 26. Mol Ther. 2010. PMID: 20104209 Free PMC article. Review.
-
A new approach to gene therapy using Sleeping Beauty to genetically modify clinical-grade T cells to target CD19.Immunol Rev. 2014 Jan;257(1):181-90. doi: 10.1111/imr.12137. Immunol Rev. 2014. PMID: 24329797 Free PMC article. Review.
Cited by
-
Manufacture of T cells using the Sleeping Beauty system to enforce expression of a CD19-specific chimeric antigen receptor.Cancer Gene Ther. 2015 Mar;22(2):95-100. doi: 10.1038/cgt.2014.69. Epub 2015 Jan 16. Cancer Gene Ther. 2015. PMID: 25591810 Review.
-
Biomanufacturing for clinically advanced cell therapies.Nat Biomed Eng. 2018 Jun;2(6):362-376. doi: 10.1038/s41551-018-0246-6. Epub 2018 Jun 11. Nat Biomed Eng. 2018. PMID: 31011198 Free PMC article. Review.
-
PiggyBac-Generated CAR19-T Cells Plus Lenalidomide Cause Durable Complete Remission of Triple-Hit Refractory/Relapsed DLBCL: A Case Report.Front Immunol. 2021 May 25;12:599493. doi: 10.3389/fimmu.2021.599493. eCollection 2021. Front Immunol. 2021. PMID: 34113336 Free PMC article. Clinical Trial.
-
Current status and hurdles for CAR-T cell immune therapy.Blood Sci. 2019 Oct 21;1(2):148-155. doi: 10.1097/BS9.0000000000000025. eCollection 2019 Oct. Blood Sci. 2019. PMID: 35402809 Free PMC article.
-
Faster, cheaper, safer, T-cell engineering.J Immunother. 2013 Jan;36(1):1-2. doi: 10.1097/CJI.0b013e3182791257. J Immunother. 2013. PMID: 23211627 Free PMC article. No abstract available.
References
-
- Bachmann M.F. Wolint P. Schwarz K. Jager P. Oxenius A. Functional properties and lineage relationship of CD8+ T cell subsets identified by expression of IL-7 receptor α and CD62L. J. Immunol. 2005;175:4686–4696. - PubMed
-
- Biagi E. Marin V. Giordano Attianese G.M. Dander E. D'Amico G. Biondi A. Chimeric T-cell receptors: New challenges for targeted immunotherapy in hematologic malignancies. Haematologica. 2007;92:381–388. - PubMed
-
- Cary L.C. Goebel M. Corsaro B.G. Wang H.G. Rosen E. Fraser M.J. Transposon mutagenesis of baculoviruses: Analysis of Trichoplusia ni transposon IFP2 insertions within the FP-locus of nuclear polyhedrosis viruses. Virology. 1989;172:156–169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
