

Dephosphorylation of gamma H2A by Glc7/protein phosphatase 1 promotes recovery from inhibition of DNA replication
- PMID: 19884341
- PMCID: PMC2798293
- DOI: 10.1128/MCB.01000-09
Dephosphorylation of gamma H2A by Glc7/protein phosphatase 1 promotes recovery from inhibition of DNA replication
Abstract
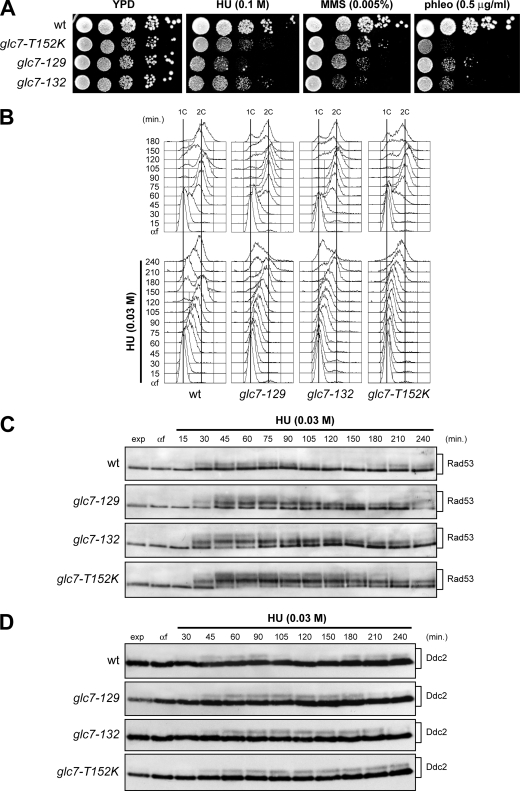
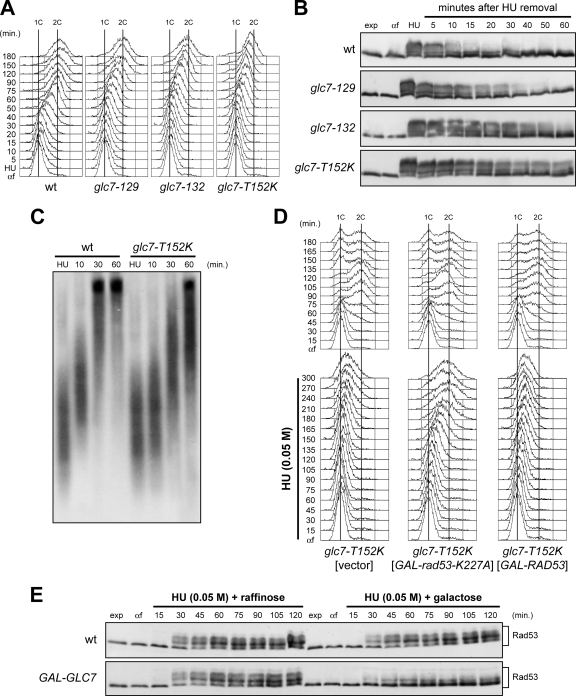
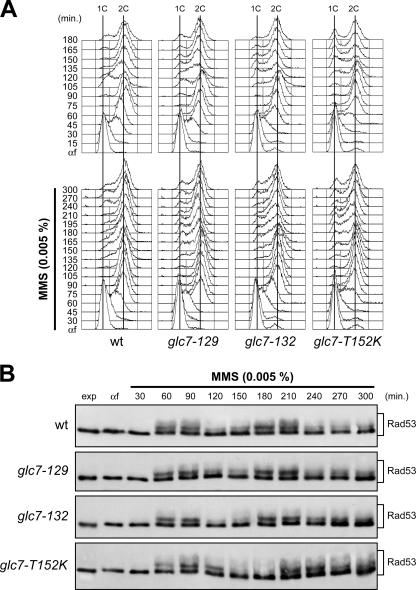
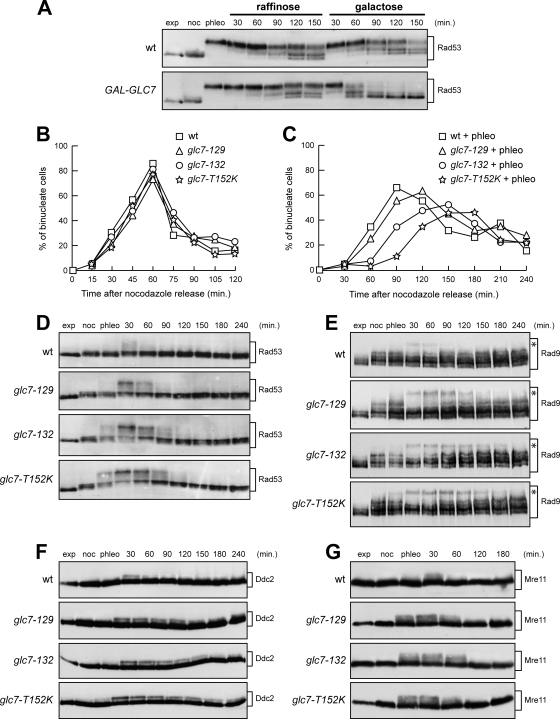
Replication fork stalling caused by deoxynucleotide depletion triggers Rad53 phosphorylation and subsequent checkpoint activation, which in turn play a crucial role in maintaining functional DNA replication forks. How cells regulate checkpoint deactivation after inhibition of DNA replication is poorly understood. Here, we show that the budding yeast protein phosphatase Glc7/protein phosphatase 1 (PP1) promotes disappearance of phosphorylated Rad53 and recovery from replication fork stalling caused by the deoxynucleoside triphosphate (dNTP) synthesis inhibitor hydroxyurea (HU). Glc7 is also required for recovery from a double-strand break-induced checkpoint, while it is dispensable for checkpoint inactivation during methylmethane sulfonate exposure, which instead requires the protein phosphatases Pph3, Ptc2, and Ptc3. Furthermore, Glc7 counteracts in vivo histone H2A phosphorylation on serine 129 (gamma H2A) and dephosphorylates gamma H2A in vitro. Finally, the replication recovery defects of HU-treated glc7 mutants are partially rescued by Rad53 inactivation or lack of gamma H2A formation, and the latter also counteracts hyperphosphorylated Rad53 accumulation. We therefore propose that Glc7 activity promotes recovery from replication fork stalling caused by dNTP depletion and that gamma H2A dephosphorylation is a critical Glc7 function in this process.
Figures
Similar articles
-
Pph3-Psy2 is a phosphatase complex required for Rad53 dephosphorylation and replication fork restart during recovery from DNA damage.Proc Natl Acad Sci U S A. 2007 May 29;104(22):9290-5. doi: 10.1073/pnas.0703252104. Epub 2007 May 21. Proc Natl Acad Sci U S A. 2007. PMID: 17517611 Free PMC article.
-
Rad53 regulates replication fork restart after DNA damage in Saccharomyces cerevisiae.Genes Dev. 2008 Jul 15;22(14):1906-20. doi: 10.1101/gad.1660408. Genes Dev. 2008. PMID: 18628397 Free PMC article.
-
Distinct phosphatases mediate the deactivation of the DNA damage checkpoint kinase Rad53.J Biol Chem. 2008 Jun 20;283(25):17123-30. doi: 10.1074/jbc.M801402200. Epub 2008 Apr 25. J Biol Chem. 2008. PMID: 18441009
-
The S-phase checkpoint: targeting the replication fork.Biol Cell. 2009 Aug 19;101(11):617-27. doi: 10.1042/BC20090053. Biol Cell. 2009. PMID: 19686094 Review.
-
Phosphatases, DNA damage checkpoints and checkpoint deactivation.Cell Cycle. 2007 Dec 15;6(24):3058-64. doi: 10.4161/cc.6.24.5100. Epub 2007 Sep 20. Cell Cycle. 2007. PMID: 18075314 Review.
Cited by
-
Yeast PP4 interacts with ATR homolog Ddc2-Mec1 and regulates checkpoint signaling.Mol Cell. 2015 Jan 22;57(2):273-89. doi: 10.1016/j.molcel.2014.11.016. Epub 2014 Dec 18. Mol Cell. 2015. PMID: 25533186 Free PMC article.
-
DNA damage checkpoint and repair: From the budding yeast Saccharomyces cerevisiae to the pathogenic fungus Candida albicans.Comput Struct Biotechnol J. 2021 Nov 25;19:6343-6354. doi: 10.1016/j.csbj.2021.11.033. eCollection 2021. Comput Struct Biotechnol J. 2021. PMID: 34938410 Free PMC article. Review.
-
Loss of H3 K79 trimethylation leads to suppression of Rtt107-dependent DNA damage sensitivity through the translesion synthesis pathway.J Biol Chem. 2010 Nov 5;285(45):35113-22. doi: 10.1074/jbc.M110.116855. Epub 2010 Sep 1. J Biol Chem. 2010. PMID: 20810656 Free PMC article.
-
Ser/Thr protein phosphatases in fungi: structure, regulation and function.Microb Cell. 2019 Apr 24;6(5):217-256. doi: 10.15698/mic2019.05.677. Microb Cell. 2019. PMID: 31114794 Free PMC article. Review.
-
Dissecting Nucleosome Function with a Comprehensive Histone H2A and H2B Mutant Library.G3 (Bethesda). 2017 Dec 4;7(12):3857-3866. doi: 10.1534/g3.117.300252. G3 (Bethesda). 2017. PMID: 29038170 Free PMC article.
References
-
- Altaf, M., A. Auger, M. Covic, and J. Côté. 2009. Connection between histone H2A variants and chromatin remodeling complexes. Biochem. Cell Biol. 87:35-50. - PubMed
-
- Bailis, J. M., and G. S. Roeder. 2000. Pachytene exit is controlled by reversal of Mek1-dependent phosphorylation. Cell 101:211-221. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases