

KIR2DS4 is a product of gene conversion with KIR3DL2 that introduced specificity for HLA-A*11 while diminishing avidity for HLA-C
- PMID: 19858347
- PMCID: PMC2768862
- DOI: 10.1084/jem.20091010
KIR2DS4 is a product of gene conversion with KIR3DL2 that introduced specificity for HLA-A*11 while diminishing avidity for HLA-C
Abstract
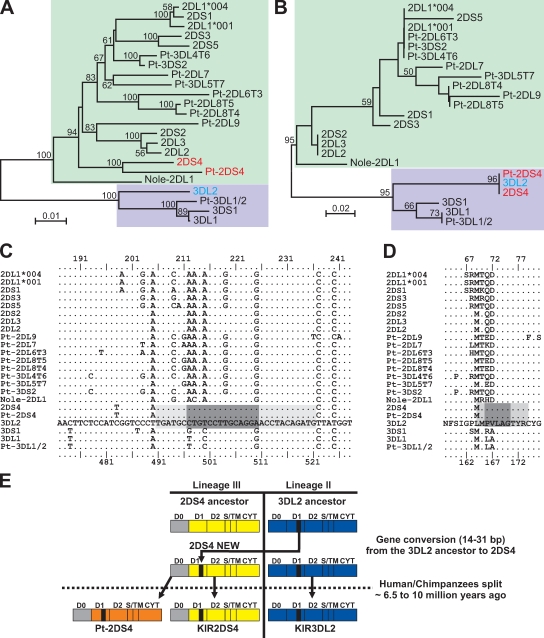
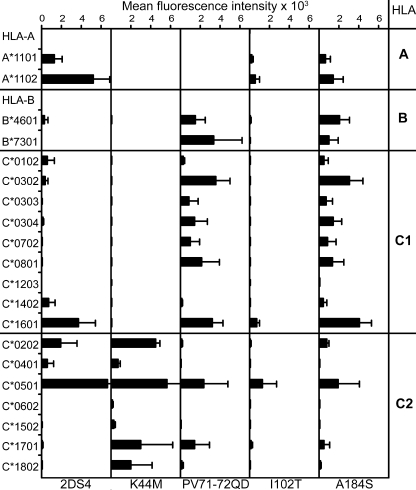
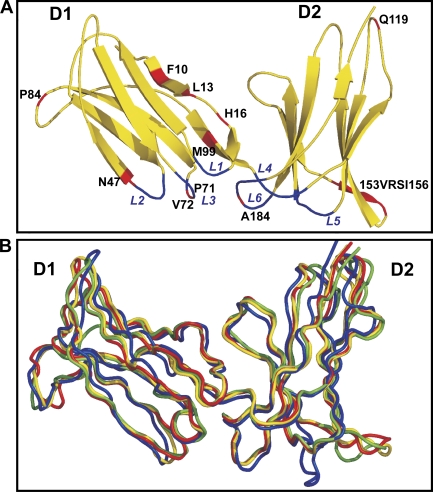
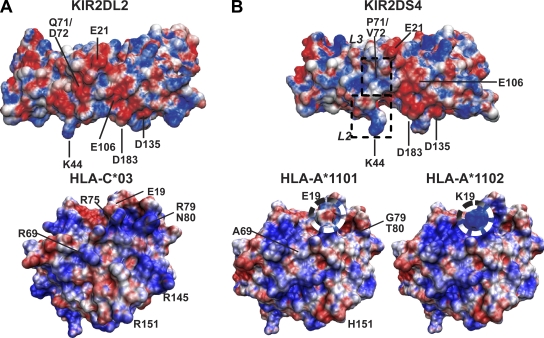
Human killer cell immunoglobulin-like receptors (KIRs) are distinguished by expansion of activating KIR2DS, whose ligands and functions remain poorly understood. The oldest, most prevalent KIR2DS is KIR2DS4, which is represented by a variable balance between "full-length" and "deleted" forms. We find that full-length 2DS4 is a human histocompatibility leukocyte antigen (HLA) class I receptor that binds specifically to subsets of C1+ and C2+ HLA-C and to HLA-A*11, whereas deleted 2DS4 is nonfunctional. Activation of 2DS4+ NKL cells was achieved with A*1102 as ligand, which differs from A*1101 by unique substitution of lysine 19 for glutamate, but not with A*1101 or HLA-C. Distinguishing KIR2DS4 from other KIR2DS is the proline-valine motif at positions 71-72, which is shared with KIR3DL2 and was introduced by gene conversion before separation of the human and chimpanzee lineages. Site-directed swap mutagenesis shows that these two residues are largely responsible for the unique HLA class I specificity of KIR2DS4. Determination of the crystallographic structure of KIR2DS4 shows two major differences from KIR2DL: displacement of contact loop L2 and altered bonding potential because of the substitutions at positions 71 and 72. Correlation between the worldwide distributions of functional KIR2DS4 and HLA-A*11 points to the physiological importance of their mutual interaction.
Figures




Similar articles
-
KIR specificity and avidity of standard and unusual C1, C2, Bw4, Bw6 and A3/11 amino acid motifs at entire HLA:KIR interface between NK and target cells, the functional and evolutionary classification of HLA class I molecules.Int J Immunogenet. 2019 Aug;46(4):217-231. doi: 10.1111/iji.12433. Epub 2019 Jun 18. Int J Immunogenet. 2019. PMID: 31210416 Review.
-
Human NK cell receptor KIR2DS4 detects a conserved bacterial epitope presented by HLA-C.Proc Natl Acad Sci U S A. 2019 Jun 25;116(26):12964-12973. doi: 10.1073/pnas.1903781116. Epub 2019 May 28. Proc Natl Acad Sci U S A. 2019. PMID: 31138701 Free PMC article.
-
Coevolution of killer cell Ig-like receptors with HLA-C to become the major variable regulators of human NK cells.J Immunol. 2010 Oct 1;185(7):4238-51. doi: 10.4049/jimmunol.1001494. Epub 2010 Aug 30. J Immunol. 2010. PMID: 20805421 Free PMC article.
-
Humans differ from other hominids in lacking an activating NK cell receptor that recognizes the C1 epitope of MHC class I.J Immunol. 2010 Oct 1;185(7):4233-7. doi: 10.4049/jimmunol.1001951. Epub 2010 Aug 27. J Immunol. 2010. PMID: 20802150 Free PMC article.
-
Natural killer cell recognition of HLA class I molecules.Rev Immunogenet. 2000;2(3):433-48. Rev Immunogenet. 2000. PMID: 11256749 Review.
Cited by
-
Donor killer immunoglobulin-like receptor genes and reactivation of cytomegalovirus after HLA-matched hematopoietic stem-cell transplantation: HLA-C allotype is an essential cofactor.Front Immunol. 2013 Feb 21;4:36. doi: 10.3389/fimmu.2013.00036. eCollection 2013. Front Immunol. 2013. PMID: 23440333 Free PMC article.
-
HLA-C*04:01 Affects HLA Class I Heterozygosity and Predicted Affinity to SARS-CoV-2 Peptides, and in Combination With Age and Sex of Armenian Patients Contributes to COVID-19 Severity.Front Immunol. 2022 Feb 3;13:769900. doi: 10.3389/fimmu.2022.769900. eCollection 2022. Front Immunol. 2022. PMID: 35185875 Free PMC article.
-
The structural basis of ligand recognition by natural killer cell receptors.J Biomed Biotechnol. 2011;2011:203628. doi: 10.1155/2011/203628. Epub 2011 May 18. J Biomed Biotechnol. 2011. PMID: 21629745 Free PMC article. Review.
-
HLA-DPB1*13:01 associates with enhanced, and KIR2DS4*001 with diminished protection from developing severe COVID-19.HLA. 2024 Jan;103(1):e15251. doi: 10.1111/tan.15251. Epub 2023 Oct 18. HLA. 2024. PMID: 37850268 Free PMC article.
-
Killer Ig-Like Receptors (KIRs): Their Role in NK Cell Modulation and Developments Leading to Their Clinical Exploitation.Front Immunol. 2019 May 28;10:1179. doi: 10.3389/fimmu.2019.01179. eCollection 2019. Front Immunol. 2019. PMID: 31231370 Free PMC article. Review.
References
-
- Adams P.D., Grosse-Kunstleve R.W., Hung L.W., Ioerger T.R., McCoy A.J., Moriarty N.W., Read R.J., Sacchettini J.C., Sauter N.K., Terwilliger T.C. 2002. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 58:1948–1954 10.1107/S0907444902016657 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
