

Circadian amplitude of cryptochrome 1 is modulated by mRNA stability regulation via cytoplasmic hnRNP D oscillation
- PMID: 19858287
- PMCID: PMC2798294
- DOI: 10.1128/MCB.01154-09
Circadian amplitude of cryptochrome 1 is modulated by mRNA stability regulation via cytoplasmic hnRNP D oscillation
Abstract
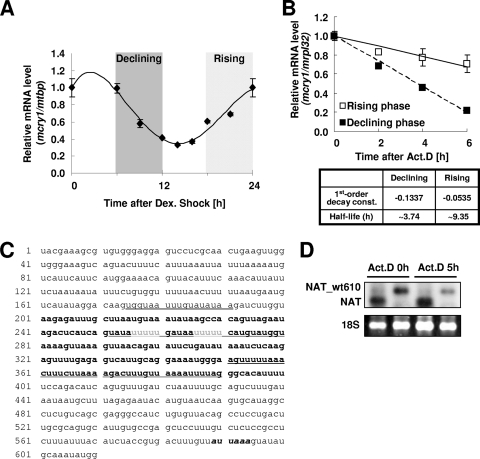
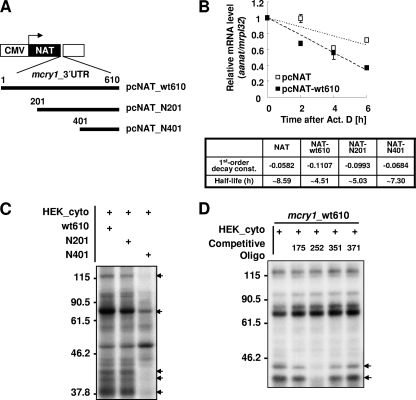
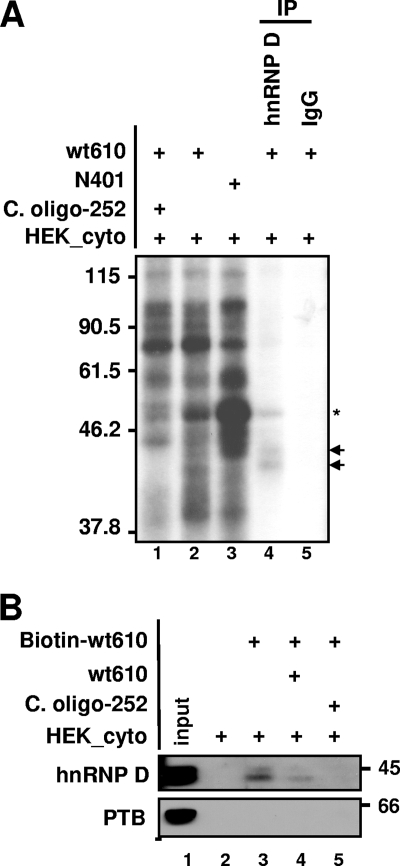
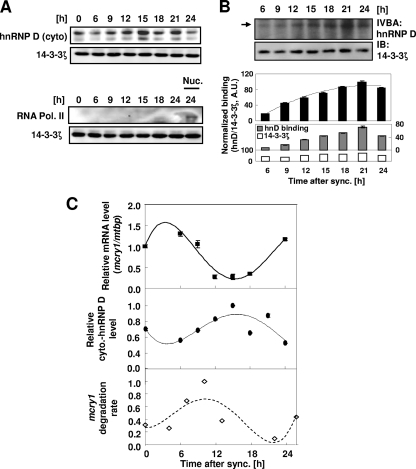
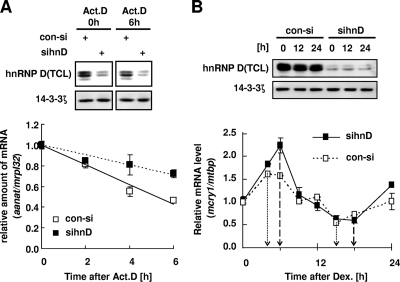
The mammalian circadian rhythm is observed not only at the suprachiasmatic nucleus, a master pacemaker, but also throughout the peripheral tissues. Its conserved molecular basis has been thought to consist of intracellular transcriptional feedback loops of key clock genes. However, little is known about posttranscriptional regulation of these genes. In the present study, we investigated the role of the 3'-untranslated region (3'UTR) of the mouse cryptochrome 1 (mcry1) gene at the posttranscriptional level. Mature mcry1 mRNA has a 610-nucleotide 3'UTR and mediates its own degradation. The middle part of the 3'UTR contains a destabilizing cis-acting element. The deletion of this element led to a dramatic increase in mRNA stability, and heterogeneous nuclear ribonucleoprotein D (hnRNP D) was identified as an RNA binding protein responsible for this effect. Cytoplasmic hnRNP D levels displayed a pattern that was reciprocal to the mcry1 oscillation. Knockdown of hnRNP D stabilized mcry1 mRNA and resulted in enhancement of the oscillation amplitude and a slight delay of the phase. Our results suggest that hnRNP D plays a role as a fine regulator contributing to the mcry1 mRNA turnover rate and the modulation of circadian rhythm.
Figures
Similar articles
-
MicroRNA-185 oscillation controls circadian amplitude of mouse Cryptochrome 1 via translational regulation.Mol Biol Cell. 2013 Jul;24(14):2248-55. doi: 10.1091/mbc.E12-12-0849. Epub 2013 May 22. Mol Biol Cell. 2013. PMID: 23699394 Free PMC article.
-
Rhythmic control of mRNA stability modulates circadian amplitude of mouse Period3 mRNA.J Neurochem. 2015 Mar;132(6):642-56. doi: 10.1111/jnc.13027. Epub 2015 Mar 9. J Neurochem. 2015. PMID: 25581122
-
AUF1 contributes to Cryptochrome1 mRNA degradation and rhythmic translation.Nucleic Acids Res. 2014 Apr;42(6):3590-606. doi: 10.1093/nar/gkt1379. Epub 2014 Jan 13. Nucleic Acids Res. 2014. PMID: 24423872 Free PMC article.
-
A position-specific 3'UTR sequence that accelerates mRNA decay.RNA Biol. 2016 Nov;13(11):1075-1077. doi: 10.1080/15476286.2016.1225645. Epub 2016 Aug 26. RNA Biol. 2016. PMID: 27565004 Free PMC article. Review.
-
Heterogeneous nuclear ribonucleoproteins (hnRNPs) in cellular processes: Focus on hnRNP E1's multifunctional regulatory roles.RNA. 2010 Aug;16(8):1449-62. doi: 10.1261/rna.2254110. Epub 2010 Jun 28. RNA. 2010. PMID: 20584894 Free PMC article. Review.
Cited by
-
Elucidating the in vivo interactome of HIV-1 RNA by hybridization capture and mass spectrometry.Sci Rep. 2017 Dec 5;7(1):16965. doi: 10.1038/s41598-017-16793-5. Sci Rep. 2017. PMID: 29208937 Free PMC article.
-
New insights into non-transcriptional regulation of mammalian core clock proteins.J Cell Sci. 2020 Sep 15;133(18):jcs241174. doi: 10.1242/jcs.241174. J Cell Sci. 2020. PMID: 32934011 Free PMC article. Review.
-
RNA around the clock - regulation at the RNA level in biological timing.Front Plant Sci. 2015 May 5;6:311. doi: 10.3389/fpls.2015.00311. eCollection 2015. Front Plant Sci. 2015. PMID: 25999975 Free PMC article. Review.
-
Circadian mRNA expression: insights from modeling and transcriptomics.Cell Mol Life Sci. 2016 Feb;73(3):497-521. doi: 10.1007/s00018-015-2072-2. Epub 2015 Oct 26. Cell Mol Life Sci. 2016. PMID: 26496725 Free PMC article. Review.
-
Introduction to the Clock System.Adv Exp Med Biol. 2021;1344:3-20. doi: 10.1007/978-3-030-81147-1_1. Adv Exp Med Biol. 2021. PMID: 34773223
References
-
- Balsalobre, A., S. A. Brown, L. Marcacci, F. Tronche, C. Kellendonk, H. M. Reichardt, G. Schutz, and U. Schibler. 2000. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 289:2344-2347. - PubMed
-
- Bevilacqua, A., M. C. Ceriani, S. Capaccioli, and A. Nicolin. 2003. Post-transcriptional regulation of gene expression by degradation of messenger RNAs. J. Cell Physiol. 195:356-372. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources