

Signals of growth regulation in bacteria
- PMID: 19854099
- PMCID: PMC2789310
- DOI: 10.1016/j.mib.2009.09.006
Signals of growth regulation in bacteria
Abstract
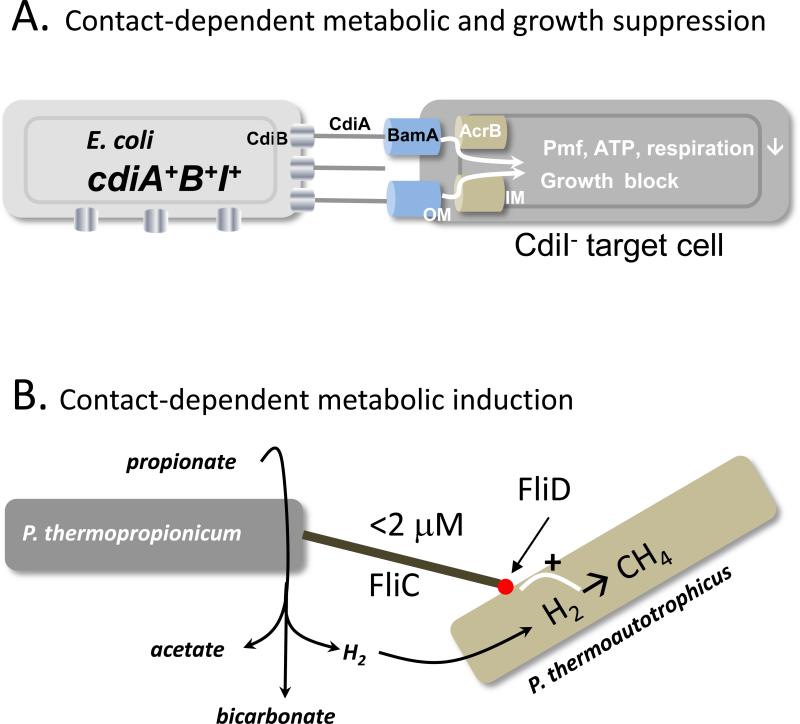
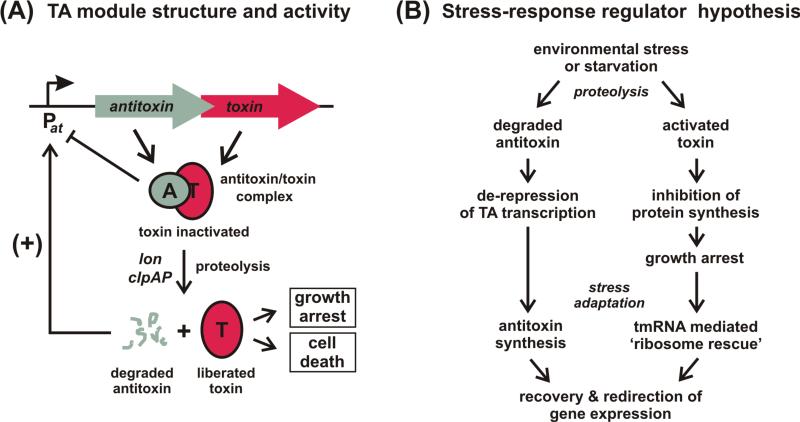
A fundamental characteristic of cells is their ability to regulate growth in response to changing environmental conditions. This review focuses on recent progress toward understanding the mechanisms by which bacterial growth is regulated. These phenomena include the 'viable but not culturable' (VBNC) state, in which bacterial growth becomes conditional, and 'persistence', which confers antibiotic resistance to a small fraction of bacteria in a population. Notably, at least one form of persistence appears to involve the generation of nongrowing phenotypic variants after transition through stationary phase. The possible roles of toxin-antitoxin modules in growth control are explored, as well as other mechanisms including contact-dependent growth inhibition, which regulates cellular metabolism and growth through binding to an outer membrane protein receptor.
Figures
Similar articles
-
Combatting bacterial persister cells.Biotechnol Bioeng. 2016 Mar;113(3):476-83. doi: 10.1002/bit.25721. Epub 2015 Sep 3. Biotechnol Bioeng. 2016. PMID: 26264116 Review.
-
Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology.Mol Cell. 2018 Jun 7;70(5):768-784. doi: 10.1016/j.molcel.2018.01.003. Epub 2018 Feb 3. Mol Cell. 2018. PMID: 29398446 Review.
-
Recent findings on the viable but nonculturable state in pathogenic bacteria.FEMS Microbiol Rev. 2010 Jul;34(4):415-25. doi: 10.1111/j.1574-6976.2009.00200.x. Epub 2009 Nov 24. FEMS Microbiol Rev. 2010. PMID: 20059548 Review.
-
Bacterial viability and culturability.Adv Microb Physiol. 1999;41:93-137. doi: 10.1016/s0065-2911(08)60166-6. Adv Microb Physiol. 1999. PMID: 10500845 Review.
-
[Viable but non-culturable form of bacteria].Mikrobiyol Bul. 2007 Jul;41(3):477-84. Mikrobiyol Bul. 2007. PMID: 17933263 Review. Turkish.
Cited by
-
Insights into the virulence of oral biofilms: discoveries from proteomics.Expert Rev Proteomics. 2012 Jun;9(3):311-23. doi: 10.1586/epr.12.16. Expert Rev Proteomics. 2012. PMID: 22809209 Free PMC article. Review.
-
Separation by dielectrophoresis of dormant and nondormant bacterial cells of Mycobacterium smegmatis.Biomicrofluidics. 2010 Jun 29;4(2):022809. doi: 10.1063/1.3435335. Biomicrofluidics. 2010. PMID: 20697591 Free PMC article.
-
Three new RelE-homologous mRNA interferases of Escherichia coli differentially induced by environmental stresses.Mol Microbiol. 2010 Jan;75(2):333-48. doi: 10.1111/j.1365-2958.2009.06969.x. Epub 2009 Nov 25. Mol Microbiol. 2010. PMID: 19943910 Free PMC article.
-
The pdh operon is expressed in a subpopulation of stationary-phase bacteria and is important for survival of sugar-starved Streptococcus mutans.J Bacteriol. 2010 Sep;192(17):4395-402. doi: 10.1128/JB.00574-10. Epub 2010 Jun 25. J Bacteriol. 2010. PMID: 20581205 Free PMC article.
-
Deletion of pknG Abates Reactivation of Latent Mycobacterium tuberculosis in Mice.Antimicrob Agents Chemother. 2021 Mar 18;65(4):e02095-20. doi: 10.1128/AAC.02095-20. Print 2021 Mar 18. Antimicrob Agents Chemother. 2021. PMID: 33468473 Free PMC article.
References
-
- Xu H, Roberts N, Singleton FL, Attwell RW, Grimes DJ, Colwell RR. Survival and viability of nonculturable Escherichia coli and Vibrio cholerae in the estuarine and marine environment. Microb. Ecol. 1982;8:313–323. - PubMed
-
- Barer MR. Viable but non-culturable and dormant bacteria: time to resolve an oxymoron and a misnomer? J Med Microbiol. 1997;46:629–631. - PubMed
-
- Coutard F, Crassous P, Droguet M, Gobin E, Colwell RR, Pommepuy M, Hervio-Heath D. Recovery in culture of viable but nonculturable Vibrio parahaemolyticus: regrowth or resuscitation? Isme J. 2007;1:111–120. - PubMed
-
- Kong I-S, Bates TC, Hulsmann A, Hassan H, Smith BE, Oliver JD. Role of catalase and oxyR in the viable but nonculturable state of Vibrio vulnificus. FEMS Microbiol Ecol. 2004;50:133–142. - PubMed
-
- Abe A, Ohashi E, Ren H, Hayashi T, Endo H. Isolation and characterization of a cold-induced nonculturable suppression mutant of Vibrio vulnificus. Microbiol Res. 2007;162:130–138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
