

The higher structure of chromatin in the LCR of the beta-globin locus changes during development
- PMID: 19781549
- PMCID: PMC2849742
- DOI: 10.1016/j.jmb.2009.09.046
The higher structure of chromatin in the LCR of the beta-globin locus changes during development
Abstract
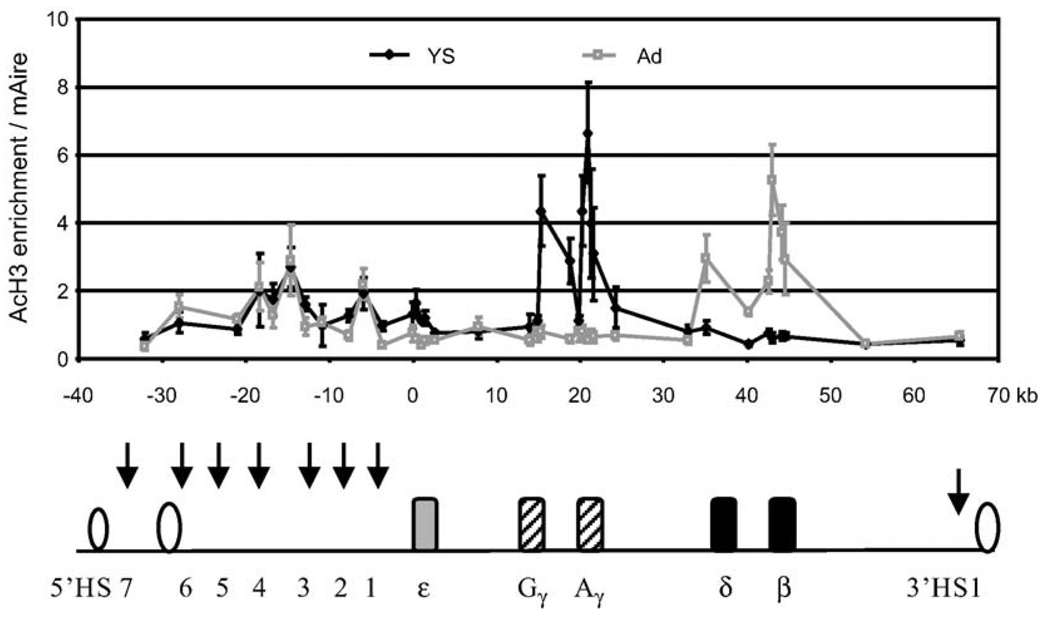
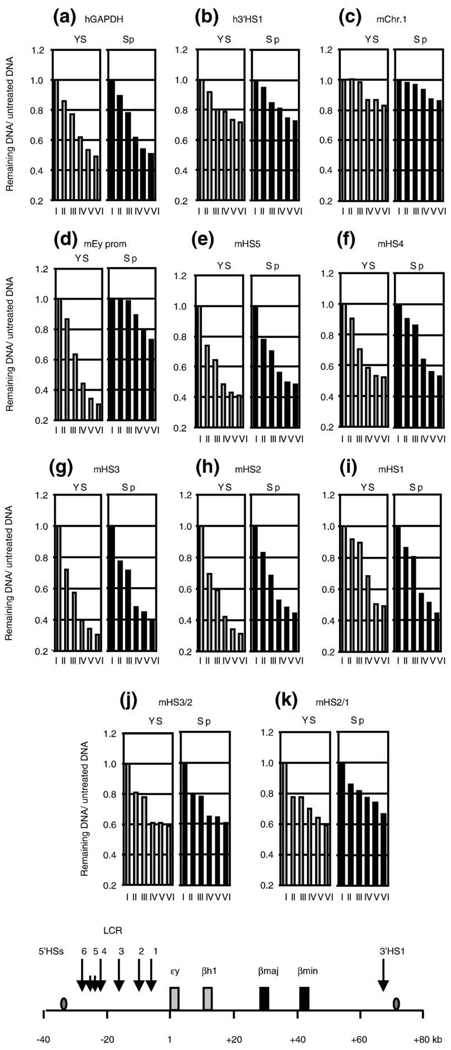
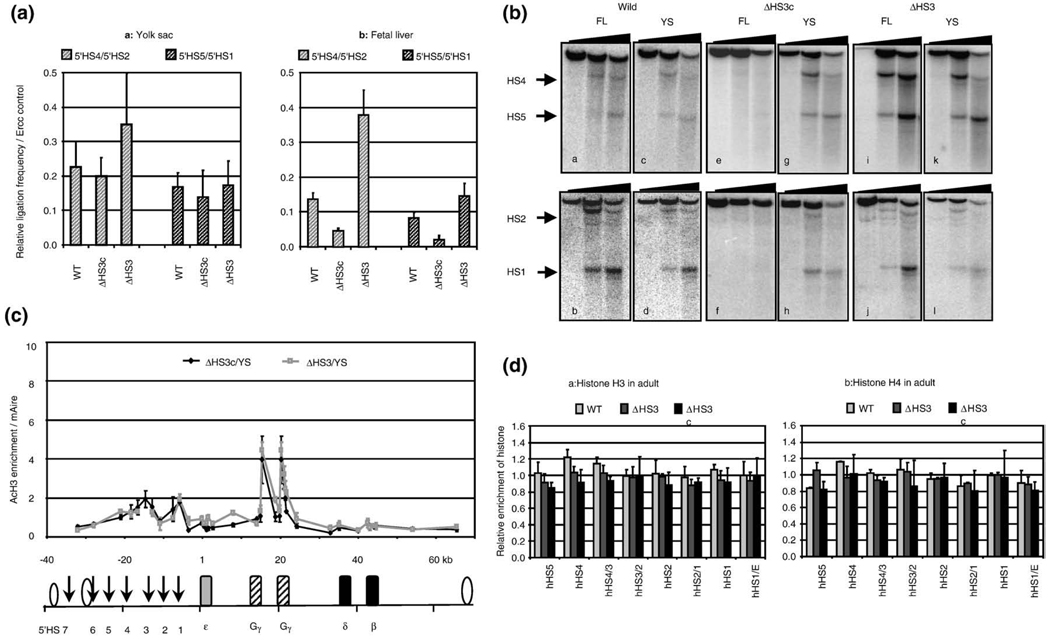
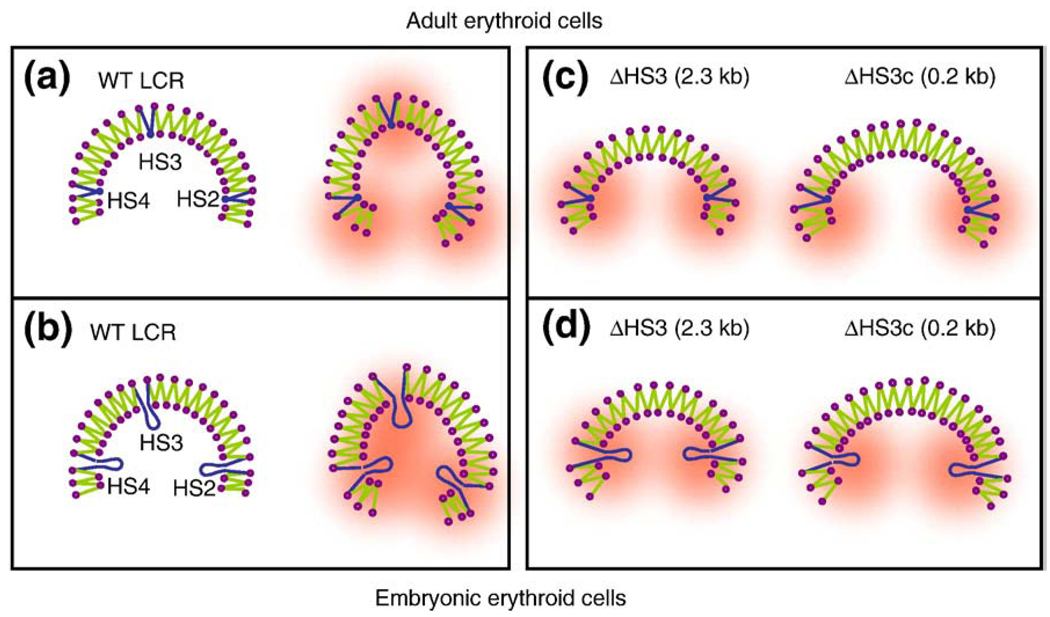
The beta-globin locus control region (LCR) is able to enhance the expression of all globin genes throughout the course of development. However, the chromatin structure of the LCR at the different developmental stages is not well defined. We report DNase I and micrococcal nuclease hypersensitivity, chromatin immunoprecipitation analyses for histones H2A, H2B, H3, and H4, and 3C (chromatin conformation capture) assays of the normal and mutant beta-globin loci, which demonstrate that nucleosomes at the DNase I hypersensitive sites of the LCR could be either depleted or retained depending on the stages of development. Furthermore, MNase sensitivity and 3C assays suggest that the LCR chromatin is more open in embryonic erythroblasts than in definitive erythroblasts at the primary- and secondary-structure levels; however, the LCR chromatin is packaged more tightly in embryonic erythroblasts than in definitive erythroblasts at the tertiary chromatin level. Our study provides the first evidence that the occupancy of nucleosomes at a DNase I hypersensitive site is a developmental stage-related event and that embryonic and adult cells possess distinct chromatin structures of the LCR.
Figures


Similar articles
-
Synergistic and additive properties of the beta-globin locus control region (LCR) revealed by 5'HS3 deletion mutations: implication for LCR chromatin architecture.Mol Cell Biol. 2005 Aug;25(16):7033-41. doi: 10.1128/MCB.25.16.7033-7041.2005. Mol Cell Biol. 2005. PMID: 16055715 Free PMC article.
-
Sequential changes in chromatin structure during transcriptional activation in the beta globin LCR and its target gene.Int J Biochem Cell Biol. 2010 Sep;42(9):1517-24. doi: 10.1016/j.biocel.2010.05.009. Epub 2010 Jun 1. Int J Biochem Cell Biol. 2010. PMID: 20561915
-
Cooperativeness of the higher chromatin structure of the beta-globin locus revealed by the deletion mutations of DNase I hypersensitive site 3 of the LCR.J Mol Biol. 2007 Jan 5;365(1):31-7. doi: 10.1016/j.jmb.2006.09.072. Epub 2006 Oct 3. J Mol Biol. 2007. PMID: 17056066 Free PMC article.
-
Structural polymorphism at LCR and its role in beta-globin gene regulation.Biochimie. 2010 Sep;92(9):1199-206. doi: 10.1016/j.biochi.2010.04.020. Epub 2010 May 11. Biochimie. 2010. PMID: 20466031 Review.
-
Joining the loops: beta-globin gene regulation.IUBMB Life. 2008 Dec;60(12):824-33. doi: 10.1002/iub.129. IUBMB Life. 2008. PMID: 18767169 Review.
Cited by
-
Transcription factors KLF1 and KLF2 positively regulate embryonic and fetal beta-globin genes through direct promoter binding.J Biol Chem. 2011 Jul 15;286(28):24819-27. doi: 10.1074/jbc.M111.247536. Epub 2011 May 24. J Biol Chem. 2011. PMID: 21610079 Free PMC article.
-
Breakpoint regions of ETO gene involved in (8; 21) leukemic translocations are enriched in acetylated histone H3.J Cell Biochem. 2013 Nov;114(11):2569-76. doi: 10.1002/jcb.24605. J Cell Biochem. 2013. PMID: 23744730 Free PMC article.
-
The distinctive roles of erythroid specific activator GATA-1 and NF-E2 in transcription of the human fetal γ-globin genes.Nucleic Acids Res. 2011 Sep 1;39(16):6944-55. doi: 10.1093/nar/gkr253. Epub 2011 May 24. Nucleic Acids Res. 2011. PMID: 21609963 Free PMC article.
-
Enhancer networks revealed by correlated DNAse hypersensitivity states of enhancers.Nucleic Acids Res. 2013 Aug;41(14):6828-38. doi: 10.1093/nar/gkt374. Epub 2013 May 21. Nucleic Acids Res. 2013. PMID: 23700312 Free PMC article.
-
Efficient genome editing in hematopoietic stem cells with helper-dependent Ad5/35 vectors expressing site-specific endonucleases under microRNA regulation.Mol Ther Methods Clin Dev. 2015 Jan 14;1:14057. doi: 10.1038/mtm.2014.57. eCollection 2015. Mol Ther Methods Clin Dev. 2015. PMID: 26052525 Free PMC article.
References
-
- Grosveld F. Activation by locus control regions? Curr. Opin. Genet. Dev. 1999;9:152–157. - PubMed
-
- Felsenfeld G, Groudine M. Controlling the double helix. Nature. 2003;421:448–453. - PubMed
-
- Engel JD, Tanimoto K. Looping, linking, and chromatin activity: new insights into beta-globin locus regulation. Cell. 2000;100:499–502. - PubMed
-
- Grosveld F, van Assendelft GB, Greaves DR, Kollias G. Position-independent, high-level expression of the human beta-globin gene in transgenic mice. Cell. 1987;51:975–985. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
