

Review
doi: 10.1038/nrn2719.
Evolution of the neocortex: a perspective from developmental biology
Affiliations
- PMID: 19763105
- PMCID: PMC2913577
- DOI: 10.1038/nrn2719
Item in Clipboard
Review
Evolution of the neocortex: a perspective from developmental biology
Nat Rev Neurosci.
2009 Oct.
Abstract
The enlargement and species-specific elaboration of the cerebral neocortex during evolution holds the secret to the mental abilities of humans; however, the genetic origin and cellular mechanisms that generated the distinct evolutionary advancements are not well understood. This article describes how novelties that make us human may have been introduced during evolution, based on findings in the embryonic cerebral cortex in different mammalian species. The data on the differences in gene expression, new molecular pathways and novel cellular interactions that have led to these evolutionary advances may also provide insight into the pathogenesis and therapies for human-specific neuropsychiatric disorders.
Figures
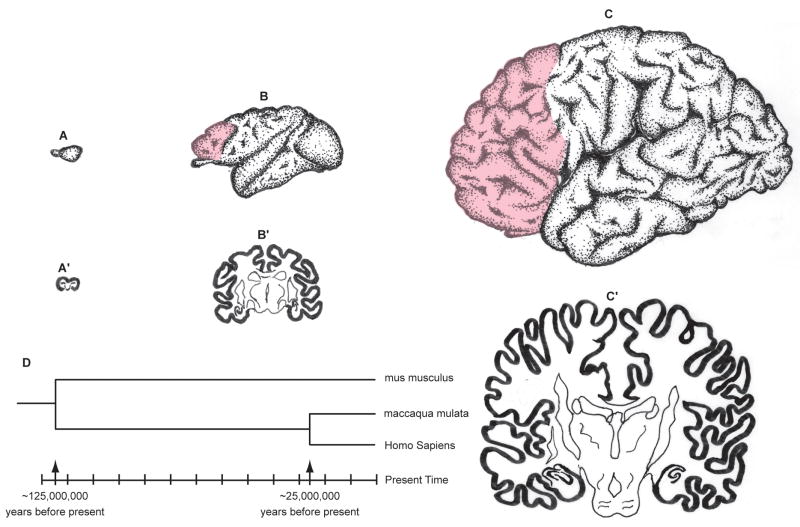
Cerebral hemispheres of the mouse (A), macaque monkey (B) and human (C) drawn at approximately the same scale to convey the overall difference in size and elaboration. The pink overlay indicates the prefrontal cortex that has no counterpart in mouse. The sections across cerebra of the same species (A′, B′, C′) illustrate the relative small increase in the thickness of the cortex compared to a large difference in surface of 1: 100: 1000 X in mouse, macaque monkey and human, respectively. D. The time-scale of phylogenetic divergence of mus musculus, maccaca mulata and homo sapiens based on the DNA sequencing data (Reviewed in REFS. 3, 4)

Figure 2A. Radial unit lineage model of cortical neurogenesis. The model illustrates how changes in the mode and the rates of cell proliferation and/or programmed cell death within the neural stem cell pool (red circles) in the ventricular zone (VZ) that divide symmetrically (SD) at early embryonic stages causes an exponential increase in the number of radial columns. That, in turn, results in surface expansion of the cerebral cortex (CC) without changes in its thickness. In contrast, similar changes in proliferation kinetics occurring in the founder cells (blue circles), which divide asymmetrically (AsD), cause a linear increase in the number of neurons within radial columns without a change in the cortical surface area. Based on the radial unit hypothesis,. Reproduced from REF. 140). Figure 2B. The model radial neuronal migration which underlies columnar organization based on REF. ,, reproduced from REF.. The cohorts of neurons generated in the ventricular zone (VZ) traverse the intermediate (IZ) and subplate zones (SP) containing “waiting” afferents from several sources (CC, TR, MB, MA) and finally pass through the earlier generated deep layers before settling in at the interface between cortical plate (CP) and marginal zone (MZ). The positional information of the neurons in the VZ and corresponding protomap within the SP and CP is preserved during cortical expansion by transient radial glial scaffolding. You can access the animated version (Rakic/Leydon model) by clicking at: http://rakiclab.med.yale.edu/RadialMigration.html

Figure 2A. Radial unit lineage model of cortical neurogenesis. The model illustrates how changes in the mode and the rates of cell proliferation and/or programmed cell death within the neural stem cell pool (red circles) in the ventricular zone (VZ) that divide symmetrically (SD) at early embryonic stages causes an exponential increase in the number of radial columns. That, in turn, results in surface expansion of the cerebral cortex (CC) without changes in its thickness. In contrast, similar changes in proliferation kinetics occurring in the founder cells (blue circles), which divide asymmetrically (AsD), cause a linear increase in the number of neurons within radial columns without a change in the cortical surface area. Based on the radial unit hypothesis,. Reproduced from REF. 140). Figure 2B. The model radial neuronal migration which underlies columnar organization based on REF. ,, reproduced from REF.. The cohorts of neurons generated in the ventricular zone (VZ) traverse the intermediate (IZ) and subplate zones (SP) containing “waiting” afferents from several sources (CC, TR, MB, MA) and finally pass through the earlier generated deep layers before settling in at the interface between cortical plate (CP) and marginal zone (MZ). The positional information of the neurons in the VZ and corresponding protomap within the SP and CP is preserved during cortical expansion by transient radial glial scaffolding. You can access the animated version (Rakic/Leydon model) by clicking at: http://rakiclab.med.yale.edu/RadialMigration.html
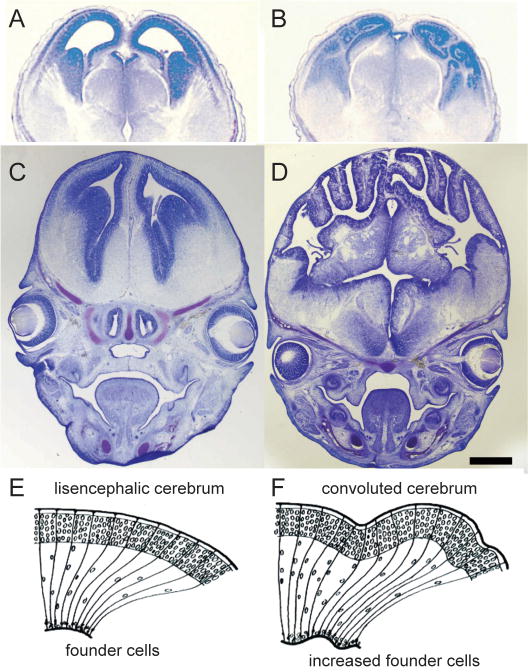
Enlargement of cortical surface by decrease in programmed cell death or increase in proliferation. Example of wildtype mouse (A) and a littermate in which both copies of Caspase 9 were deleted, later leading to expansion of the progenitor pool, increased number of radial columns, and convoluted cortical surface (B). (C, D), Enlarged brains and heads of β-catenin transgenic mice with horizontal expansion of precursor population. Mid-coronal section through the forebrain stained with cresyl violet of an embryonic day 15.5 wild-type littermate control (C) and comparable section of a transgenic animal (D) expressing a Δ90β-catenin–GFP fusion protein in neural precursors. The forebrain of transgenic animals is enlarged overall, with increased surface area and folding of the epithelial surface. Drawings in E and F are the models of how an increased number of progenitors can be distributed via radial glial scaffolding to form a sheet, rather than a lump of postmitotic neurons. A and B. reproduced from REF. ; C and D, from REF., with permission.
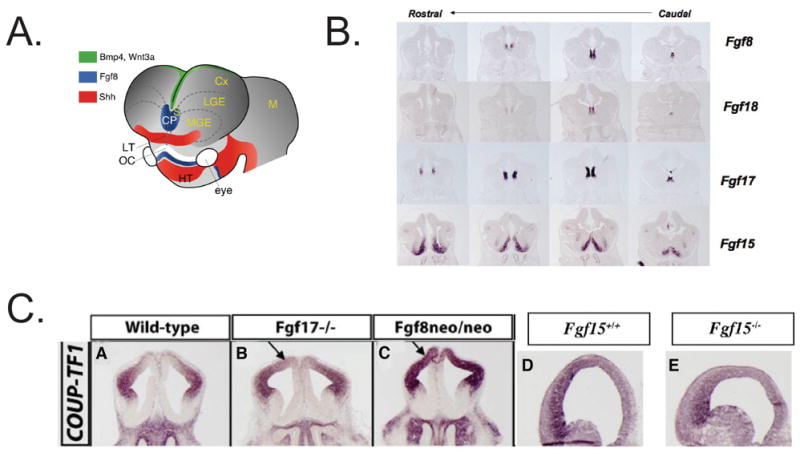
Control of arealization of the frontal cortex by Fgf expression. A. Patterning centers in the forebrain from REF . Fgf expression is in the rostral patterning center (also called commissural plate, CP). B. Fgf expression in the rostral patterning center in coronal sections of an E12.5 mouse telencephalon from REF. C. Transcription factors expressed in the E12.5 rostral telencephalon; arrows show approximate boundaries of their expression in the anlage of the frontal cortex CX: Cortex; LGE: lateral ganglionic eminence; SE: septum. Dotted lines: cortical/subcortical boundary. (Faedo and Rubenstein, unpublished). D. Repression of COUP-TF1 expression by Fgf17 (Fgf17−/− null mutant) (B), Fgf8 (hypomorphic neo/neo mutant) (C). Promotion of COUP-TF1 expression by Fgf15 (Fgf15−/− null mutant) (E). Coronal sections of E12.5 telencephalon from REFS. ,.
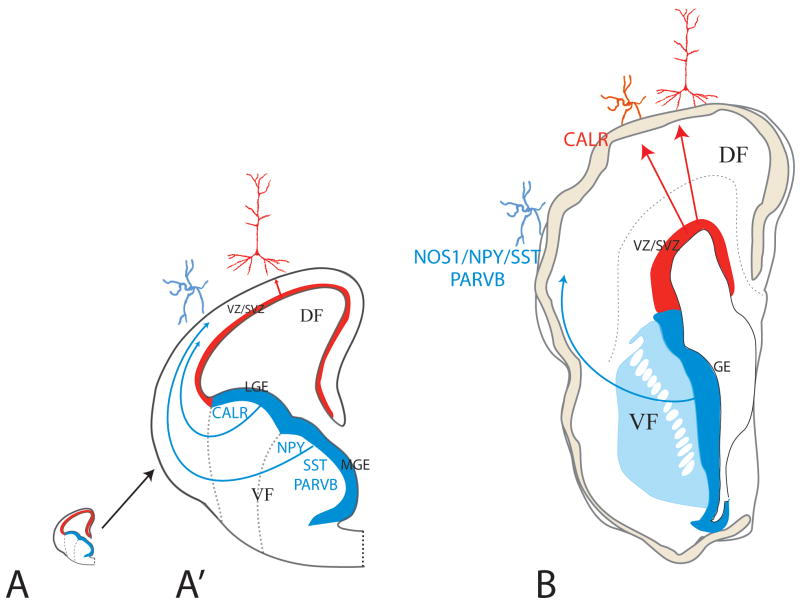
The schematic illustration of half of a cross-section through fetal rodent (A) and human (B) forebrains at the peak of corticogenesis at approximately same scale (A′ is an enlargement of the mouse forebrain to render the drawing legible). In rodents (A′), the main source of interneurons is the ganglionic eminence (GE) of the ventral telencephalon, which then migrate tangentially to the neocortex in the dorsal telencephalon. In contrast, interneurons in the human forebrain originate both in the GE as well as locally in the ventricular zone/subventricular zone (VZ/SVZ) of the dorsal telencephalon subjacent to the neocortex. Interneurons originating from the cortical VZ/SVZ and GE express different set of markers. (Figure based on REF. (mouse) and REFS. , (human).

A. Neocortical areas of the human fetal cerebral hemispheres examined by Johnson et al.(2009) included four distinct subdivisions of the prefrontal cortex (PFC) (blue): orbital, medial, dorsolateral and ventrolateral (Brodman’s areas: 9, 13, 32,44, 45) and perisylvian areas (1–3,4,40,41,42) (yellow). In addition to the ventrolateral PFC, which encompasses the prospective Broca’s area, two other speech and language-related areas were assayed: parietal and temporal association cortices, which encompass the prospective Geschwind’s and Wernicke’s areas, respectively. B-C. Examples of different clusters of correlated gene expression patterns in the PFC and perisylvian (C) cortical areas (Adapted from REF. 124).
Similar articles
-
Cajal-Retzius cells and the development of the neocortex.Trends Neurosci. 1998 Feb;21(2):64-71. doi: 10.1016/s0166-2236(97)01164-8. Trends Neurosci. 1998. PMID: 9498301 Review.
-
Neocortical neurogenesis is not really "neo": a new evolutionary model derived from a comparative study of chick pallial development.Dev Growth Differ. 2013 Jan;55(1):173-87. doi: 10.1111/dgd.12020. Epub 2012 Dec 12. Dev Growth Differ. 2013. PMID: 23230908 Review.
-
Origin and evolution of developmental enhancers in the mammalian neocortex.Proc Natl Acad Sci U S A. 2016 May 10;113(19):E2617-26. doi: 10.1073/pnas.1603718113. Epub 2016 Apr 25. Proc Natl Acad Sci U S A. 2016. PMID: 27114548 Free PMC article.
-
How unique is the human neocortex?Development. 2014 Jan;141(1):11-6. doi: 10.1242/dev.101279. Development. 2014. PMID: 24346696
-
Human neocortical development: the importance of embryonic and early fetal events.Neuroscientist. 2001 Aug;7(4):303-14. doi: 10.1177/107385840100700407. Neuroscientist. 2001. PMID: 11488396 Review.
Cited by
-
General hallmarks of microRNAs in brain evolution and development.RNA Biol. 2015;12(7):701-8. doi: 10.1080/15476286.2015.1048954. RNA Biol. 2015. PMID: 26000728 Free PMC article. Review.
-
Large-scale genomics unveil polygenic architecture of human cortical surface area.Nat Commun. 2015 Jul 20;6:7549. doi: 10.1038/ncomms8549. Nat Commun. 2015. PMID: 26189703 Free PMC article.
-
Cross-site reproducibility of human cortical organoids reveals consistent cell type composition and architecture.Stem Cell Reports. 2024 Sep 10;19(9):1351-1367. doi: 10.1016/j.stemcr.2024.07.008. Epub 2024 Aug 22. Stem Cell Reports. 2024. PMID: 39178845 Free PMC article.
-
Mutant Huntingtin Drives Development of an Advantageous Brain Early in Life: Evidence in Support of Antagonistic Pleiotropy.Ann Neurol. 2024 Nov;96(5):1006-1019. doi: 10.1002/ana.27046. Epub 2024 Aug 8. Ann Neurol. 2024. PMID: 39115048
-
Lost highway(s): barriers to postnatal cortical neurogenesis and implications for brain repair.Front Cell Neurosci. 2015 Jun 16;9:216. doi: 10.3389/fncel.2015.00216. eCollection 2015. Front Cell Neurosci. 2015. PMID: 26136658 Free PMC article. Review.
References
-
- Striedter GF. Principles of Brain Evolution. Sinauer; Sunderland, MA: 2005.
-
- Northcutt RG. Evolution of the telencephalon in non-mammals. Ann Rev Neurosci. 1981;4:301–350. - PubMed
-
- Murphy WJ, Pevzner PA, O’Brian SJ. Mammalian phylogenomic comes of age. Trends Genet. 2004;20:631–9. A concise and informative review of the DNA sequencing-based time-scale of phylogenetic divergence of various mammalian species. - PubMed
-
- Preuss TM. The cognitive neuroscience of human uniqueness. In: Gazzaniga MS, editor. The Cognitive Neuroscience IV. The MIT Press; Cambridge, MA: 2009. in the press.
-
- Goffinet AM, Rakic P, editors. Mouse Brain Development. Springer-Verlag; Berlin; New York: 2000.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
