

Partial rescue of postnatal growth plate abnormalities in Ihh mutants by expression of a constitutively active PTH/PTHrP receptor
- PMID: 19761883
- PMCID: PMC2818229
- DOI: 10.1016/j.bone.2009.09.009
Partial rescue of postnatal growth plate abnormalities in Ihh mutants by expression of a constitutively active PTH/PTHrP receptor
Abstract
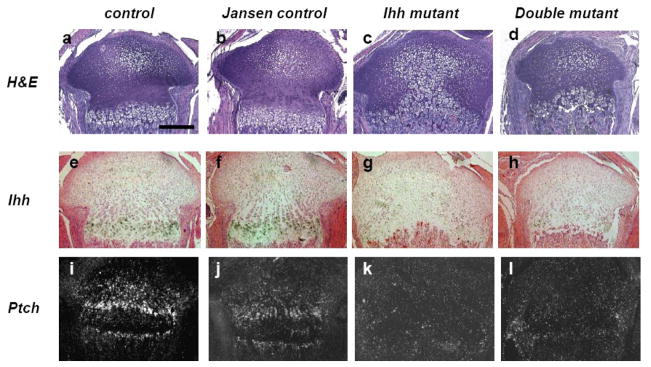
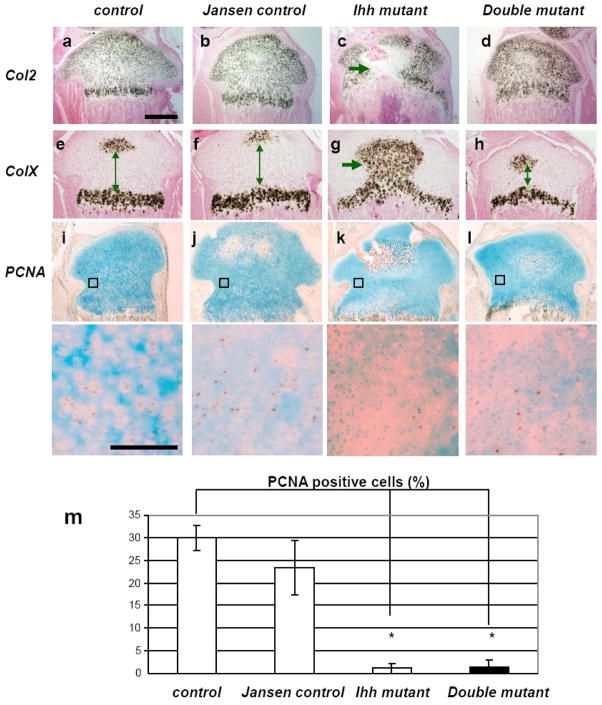
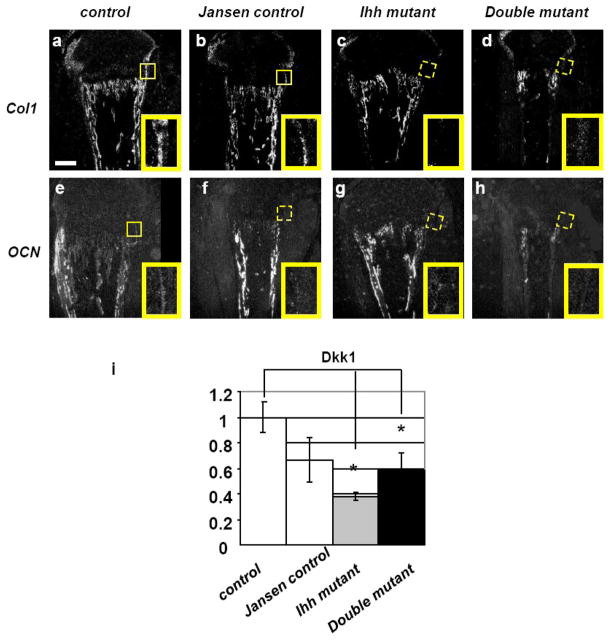
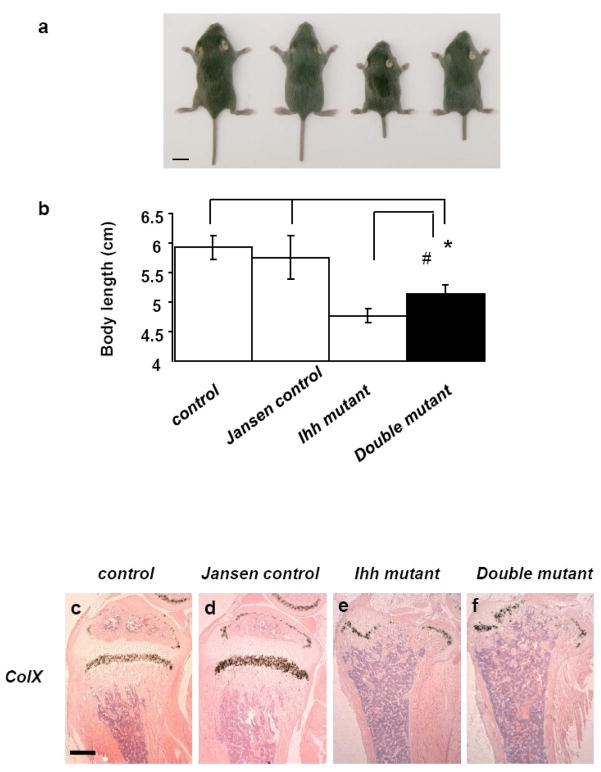
Indian hedgehog (Ihh) is essential for chondrocyte proliferation/differentiation and osteoblast differentiation during prenatal endochondral bone formation. Ihh expression in postnatal chondrocytes has a non-redundant role in maintaining a growth plate and sustaining trabecular bone after birth. Loss of Ihh in postnatal chondrocytes results in fusion of the growth plate and a decrease in trabecular bone. In order to normalize this abnormal chondrocyte phenotype and to investigate whether a putative rescue of the growth plate anomalies is sufficient to correct the severe alterations in the bone, we expressed a constitutively active PTH/PTHrP receptor (an Ihh downstream target) in the chondrocytes of Col2 alpha 1-Cre ER; Ihh(dld) mice by mating Col2 alpha 1-Cre ER; Ihh(fl/fl) mice with Col2 alpha 1-constitutively active PTH/PTHrP receptor transgenic mice (Jansen, J). Col2 alpha 1-Cre ER; Ihh(f/f); J mice were then injected with tamoxifen at P0 to generate Col2 alpha 1-Cre ER; Ihh(d/d); J mice. In contrast with the previously reported growth plate phenotype of Col2 alpha 1-Cre ER; Ihh(d/d) mice that displayed ectopic chondrocyte hypertrophy at P7, growth plates of Col2 alpha 1-Cre ER; Ihh(d/d); J double mutants were well organized, and exhibited a gene expression pattern similar to the one of control mice. However, expression of osteoblast markers and Dkk1, a Wnt signaling target, remains decreased in the bone collar of Col2 alpha 1-Cre ER; Ihh(d/d); J mice when compared to control mice despite the rescue of abnormal chondrocyte differentiation. Moreover, proliferation of chondrocytes was still significantly impaired in Col2 alpha 1-Cre ER; Ihh(d/d); J mice, and this eventually led to the fusion of the growth plate at P14. In summary, we have demonstrated that expression of a Jansen receptor in chondrocytes was able to rescue abnormal chondrocyte differentiation but not impaired chondrocyte proliferation and the bone anomalies in mice lacking the Ihh gene in chondrocytes after birth. Taken together, our findings suggest that Ihh has both PTHrP-dependent and -independent functions during postnatal endochondral bone development.
(c) 2009 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Core binding factor beta (Cbfβ) controls the balance of chondrocyte proliferation and differentiation by upregulating Indian hedgehog (Ihh) expression and inhibiting parathyroid hormone-related protein receptor (PPR) expression in postnatal cartilage and bone formation.J Bone Miner Res. 2014 Jul;29(7):1564-1574. doi: 10.1002/jbmr.2275. J Bone Miner Res. 2014. PMID: 24821091 Free PMC article.
-
Ihh and PTH1R signaling in limb mesenchyme is required for proper segmentation and subsequent formation and growth of digit bones.Bone. 2016 Feb;83:256-266. doi: 10.1016/j.bone.2015.11.017. Epub 2015 Nov 24. Bone. 2016. PMID: 26620087
-
Tamoxifen-inducible gene deletion reveals a distinct cell type associated with trabecular bone, and direct regulation of PTHrP expression and chondrocyte morphology by Ihh in growth region cartilage.Dev Biol. 2007 Aug 1;308(1):93-105. doi: 10.1016/j.ydbio.2007.05.011. Epub 2007 May 18. Dev Biol. 2007. PMID: 17560974 Free PMC article.
-
The parathyroid hormone-related protein and Indian hedgehog feedback loop in the growth plate.Novartis Found Symp. 2001;232:144-52; discussion 152-7. doi: 10.1002/0470846658.ch10. Novartis Found Symp. 2001. PMID: 11277077 Review.
-
Role of parathyroid hormone-related peptide and Indian hedgehog in skeletal development.Pediatr Nephrol. 2000 Jul;14(7):606-11. doi: 10.1007/s004670000343. Pediatr Nephrol. 2000. PMID: 10912527 Review.
Cited by
-
Developmental Regulation of the Growth Plate and Cranial Synchondrosis.J Dent Res. 2016 Oct;95(11):1221-9. doi: 10.1177/0022034516651823. Epub 2016 Jun 1. J Dent Res. 2016. PMID: 27250655 Free PMC article. Review.
-
Hedgehog Signaling in Skeletal Development: Roles of Indian Hedgehog and the Mode of Its Action.Int J Mol Sci. 2020 Sep 11;21(18):6665. doi: 10.3390/ijms21186665. Int J Mol Sci. 2020. PMID: 32933018 Free PMC article. Review.
-
Astragalus Extract Mixture HT042 Alleviates Dexamethasone-Induced Bone Growth Retardation in Rat Metatarsal Bones.Nutrients. 2024 Jul 19;16(14):2333. doi: 10.3390/nu16142333. Nutrients. 2024. PMID: 39064775 Free PMC article.
-
PTHrP treatment fails to rescue bone defects caused by Hedgehog pathway inhibition in young mice.Toxicol Pathol. 2011 Apr;39(3):478-85. doi: 10.1177/0192623311399788. Epub 2011 Mar 16. Toxicol Pathol. 2011. PMID: 21411723 Free PMC article.
-
Indian hedgehog signaling regulates transcription and expression of collagen type X via Runx2/Smads interactions.J Biol Chem. 2014 Sep 5;289(36):24898-910. doi: 10.1074/jbc.M114.570507. Epub 2014 Jul 15. J Biol Chem. 2014. PMID: 25028519 Free PMC article.
References
-
- Lanske B, Karaplis AC, Lee K, Luz A, Vortkamp A, Pirro A, Karperien M, Defize LH, Ho C, Mulligan RC, Abou-Samra AB, Juppner H, Segre GV, Kronenberg HM. PTH/PTHrP receptor in early development and Indian hedgehog-regulated bone growth. Science. 1996;273:663–6. - PubMed
-
- Vortkamp A, Lee K, Lanske B, Segre GV, Kronenberg HM, Tabin CJ. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science. 1996;273:613–22. - PubMed
-
- Long F, Chung UI, Ohba S, McMahon J, Kronenberg HM, McMahon AP. Ihh signaling is directly required for the osteoblast lineage in the endochondral skeleton. Development. 2004;131:1309–18. - PubMed
-
- Rodda SJ, McMahon AP. Distinct roles for Hedgehog and canonical Wnt signaling in specification, differentiation and maintenance of osteoblast progenitors. Development. 2006;133:3231–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
