

Pharmacological activation of guanine nucleotide exchange factors for the small GTPase Rap1 recruits high-affinity beta1 integrins as coreceptors for parvovirus B19: improved ex vivo gene transfer to human erythroid progenitor cells
- PMID: 19702438
- PMCID: PMC2829459
- DOI: 10.1089/hum.2009.123
Pharmacological activation of guanine nucleotide exchange factors for the small GTPase Rap1 recruits high-affinity beta1 integrins as coreceptors for parvovirus B19: improved ex vivo gene transfer to human erythroid progenitor cells
Abstract
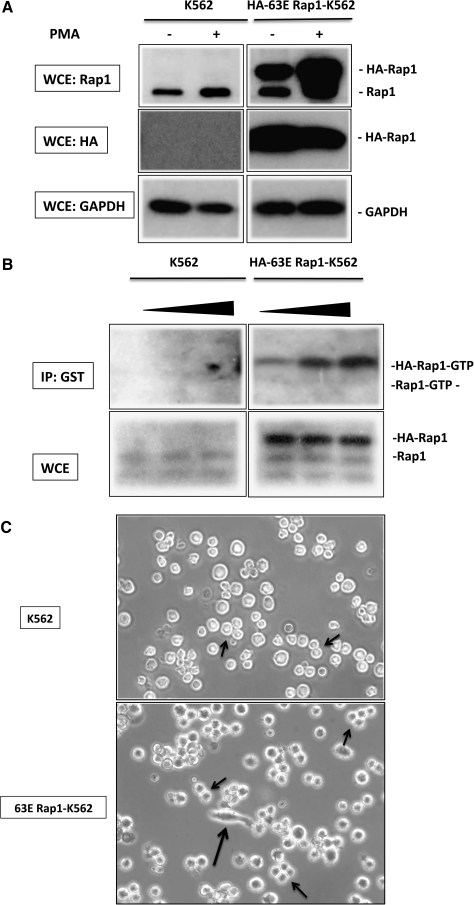
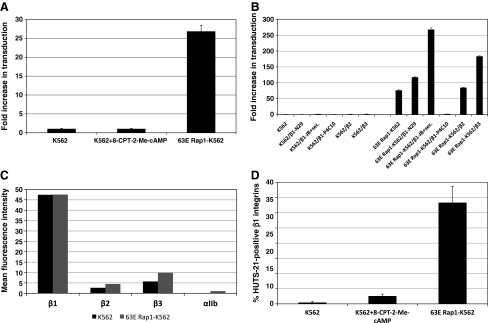
Parvovirus B19 has potential as a gene therapy vector because of its restricted tropism for human erythroid progenitor cells in the bone marrow. B19 binds to the cell surface through P antigen and we identified activated beta(1) integrins as coreceptors for internalization. Because differentiation with phorbol ester induces beta(1) integrin coreceptor activity, but cell differentiation is not desirable in gene transfer to human progenitor cells and one of the downstream effectors of phorbol esters is the small GTPase Rap1, the role of Rap1 in the recruitment of beta(1) integrins on hematopoietic cells was examined. Expression of a constitutively active Rap1 (63E) was sufficient to recruit beta(1) integrin coreceptors in erythroleukemic K562 cells by inducing high-affinity integrin conformation. A crucial role of actin polymerization in Rap1-mediated beta(1) integrin recruitment was documented by complete inhibition of the 63E Rap1 effect with low-dose cytochalasin D and by the ability of a constitutively active mutant of the actin cytoskeleton regulator Rac1 to sensitize K562 cells to the pharmacological activation of endogenous Rap1, using the Rap1 exchange factor-specific 8-pCPT-2'-O-Me-cAMP [8-(4-chlorophenylthio)-2'-O-methyladenosine-3',5'-cyclic monophosphate]. Interestingly, in primary human erythroid progenitor cells, 8-pCPT-2'-O-Me-cAMP was sufficient to significantly increase B19-mediated gene transfer, suggesting that these cells possess the cytoskeleton organization capacity required for efficient recruitment of beta(1) integrins by brief pharmacological stimulation of Rap1 GTP loading. Because 8-pCPT-2'-O-Me-cAMP has been implicated in enhanced homing of progenitor cells, these results identify a novel tool with which to optimize ex vivo B19-mediated gene transfer and potentially improve homing of transduced cells by Rap1-beta(1) integrin activation with 8-pCPT-2'-O-Me-cAMP.
Figures

Similar articles
-
Enhanced Rap1 activation and insulin secretagogue properties of an acetoxymethyl ester of an Epac-selective cyclic AMP analog in rat INS-1 cells: studies with 8-pCPT-2'-O-Me-cAMP-AM.J Biol Chem. 2009 Apr 17;284(16):10728-36. doi: 10.1074/jbc.M900166200. Epub 2009 Feb 25. J Biol Chem. 2009. PMID: 19244230 Free PMC article.
-
Alpha5beta1 integrin as a cellular coreceptor for human parvovirus B19: requirement of functional activation of beta1 integrin for viral entry.Blood. 2003 Dec 1;102(12):3927-33. doi: 10.1182/blood-2003-05-1522. Epub 2003 Aug 7. Blood. 2003. PMID: 12907437
-
Activation of Epac stimulates integrin-dependent homing of progenitor cells.Blood. 2008 Mar 1;111(5):2640-6. doi: 10.1182/blood-2007-04-086231. Epub 2007 Nov 21. Blood. 2008. PMID: 18032709
-
The role of Rap1 in integrin-mediated cell adhesion.Biochem Soc Trans. 2003 Feb;31(Pt 1):83-6. doi: 10.1042/bst0310083. Biochem Soc Trans. 2003. PMID: 12546659 Review.
-
Regulatory actions of 3',5'-cyclic adenosine monophosphate on osteoclast function: possible roles of Epac-mediated signaling.Ann N Y Acad Sci. 2018 Dec;1433(1):18-28. doi: 10.1111/nyas.13861. Epub 2018 May 30. Ann N Y Acad Sci. 2018. PMID: 29846007 Review.
Cited by
-
Advances in human B19 erythrovirus biology.J Virol. 2010 Oct;84(19):9658-65. doi: 10.1128/JVI.00684-10. Epub 2010 Jul 14. J Virol. 2010. PMID: 20631151 Free PMC article. Review.
-
The globoside receptor triggers structural changes in the B19 virus capsid that facilitate virus internalization.J Virol. 2010 Nov;84(22):11737-46. doi: 10.1128/JVI.01143-10. Epub 2010 Sep 8. J Virol. 2010. PMID: 20826697 Free PMC article.
References
-
- Arai A. Nosaka Y. Kanda E. Yamamoto K. Miyasaka N. Miura O. Rap1 is activated by erythropoietin or interleukin-3 and is involved in regulation of β1 integrin-mediated hematopoietic cell adhesion. J. Biol. Chem. 2001;276:10453–10462. - PubMed
-
- Berger G. Quarck R. Tenza D. Levy-Toledano S. De Gunzburg J. Cramer E.M. Ultrastructural localization of the small GTP-binding protein Rap1 in human platelets and megakaryocytes. Br. J. Haematol. 1994;88:372–382. - PubMed
-
- Bertoni A. Tadokoro S. Eto K. Pampori N. Parise L.V. White G.C. Shattil S.J. Relationships between Rap1b, affinity modulation of integrin αIIbβ3, and the actin cytoskeleton. J. Biol. Chem. 2002;277:25715–25721. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
