

Dynamic chromosome organization and protein localization coordinate the regulatory circuitry that drives the bacterial cell cycle
- PMID: 19687139
- PMCID: PMC7451405
- DOI: 10.1101/sqb.2009.74.005
Dynamic chromosome organization and protein localization coordinate the regulatory circuitry that drives the bacterial cell cycle
Abstract
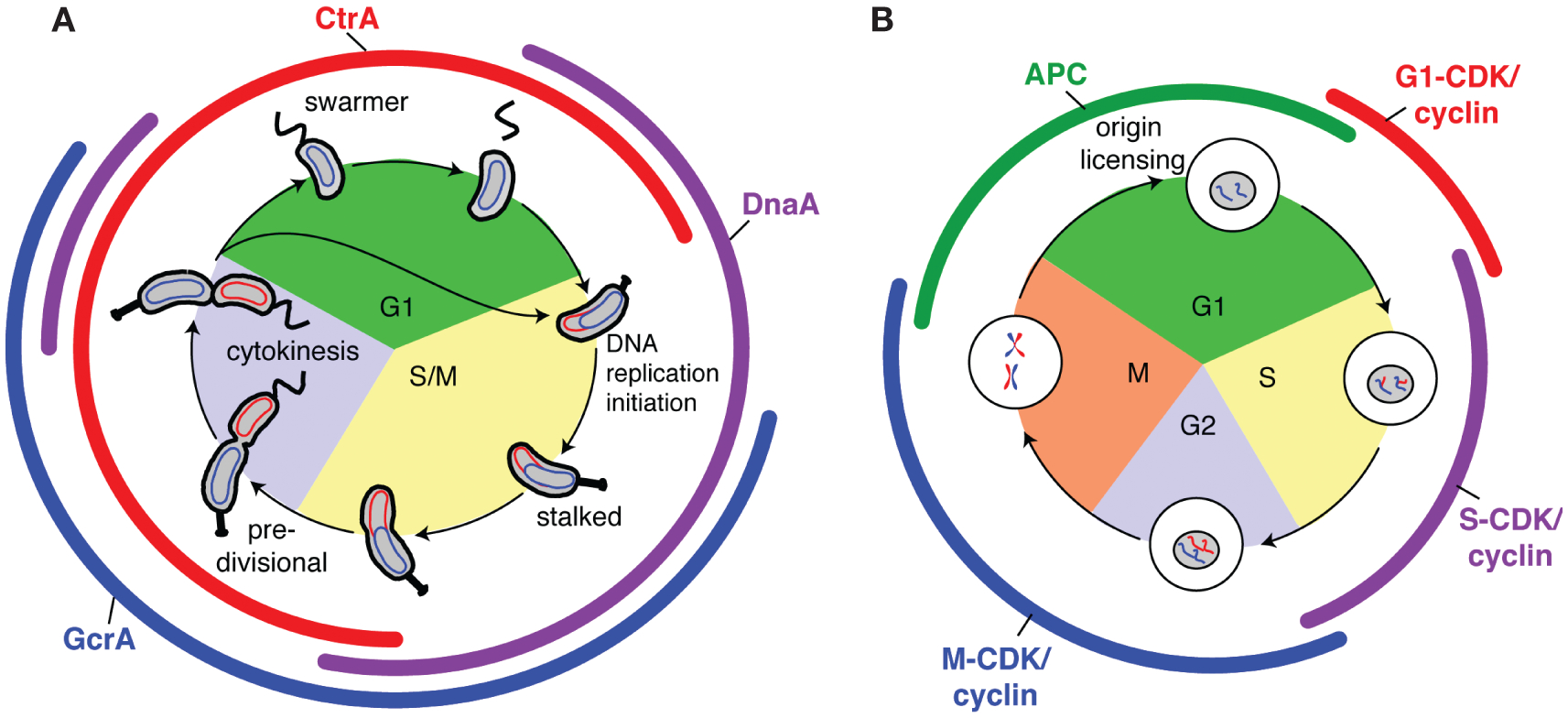
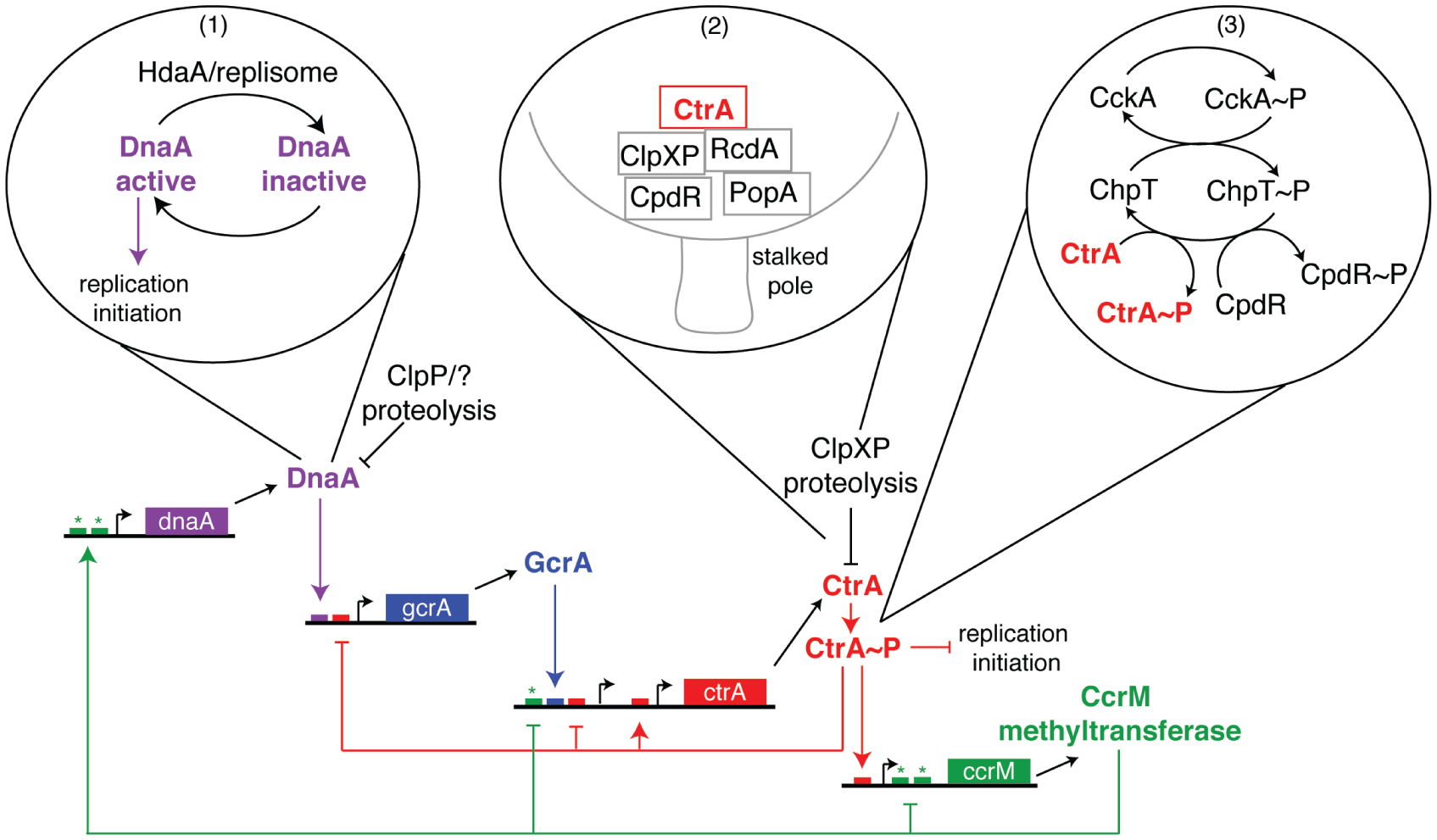
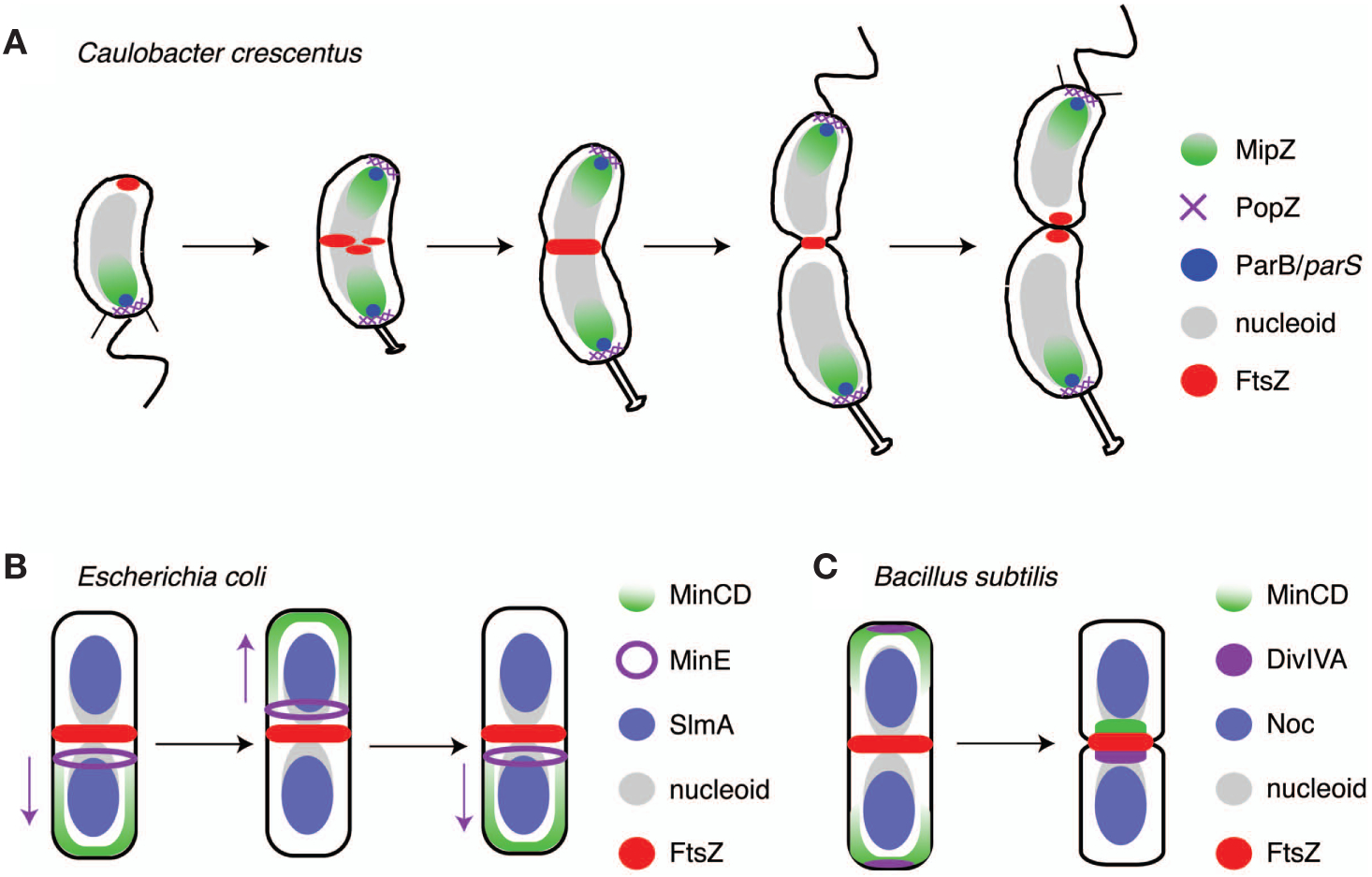
The bacterial cell has less internal structure and genetic complexity than cells of eukaryotic organisms, yet it is a highly organized system that uses both temporal and spatial cues to drive its cell cycle. Key insights into bacterial regulatory programs that orchestrate cell cycle progression have come from studies of Caulobacter crescentus, a bacterium that divides asymmetrically. Three global regulatory proteins cycle out of phase with one another and drive cell cycle progression by directly controlling the expression of 200 cell-cycle-regulated genes. Exploration of this system provided insights into the evolution of regulatory circuits and the plasticity of circuit structure. The temporal expression of the modular subsystems that implement the cell cycle and asymmetric cell division is also coordinated by differential DNA methylation, regulated proteolysis, and phosphorylation signaling cascades. This control system structure has parallels to eukaryotic cell cycle control architecture. Remarkably, the transcriptional circuitry is dependent on three-dimensional dynamic deployment of key regulatory and signaling proteins. In addition, dynamically localized DNA-binding proteins ensure that DNA segregation is coupled to the timing and cellular position of the cytokinetic ring. Comparison to other organisms reveals conservation of cell cycle regulatory logic, even if regulatory proteins, themselves, are not conserved.
Figures
Similar articles
-
Dynamic localization of proteins and DNA during a bacterial cell cycle.Nat Rev Mol Cell Biol. 2002 Mar;3(3):167-76. doi: 10.1038/nrm758. Nat Rev Mol Cell Biol. 2002. PMID: 11994737 Review.
-
Oscillating global regulators control the genetic circuit driving a bacterial cell cycle.Science. 2004 May 14;304(5673):983-7. doi: 10.1126/science.1095191. Epub 2004 Apr 15. Science. 2004. PMID: 15087506
-
Cell cycle control by oscillating regulatory proteins in Caulobacter crescentus.Bioessays. 2006 Apr;28(4):355-61. doi: 10.1002/bies.20384. Bioessays. 2006. PMID: 16547950 Review.
-
A genetic oscillator and the regulation of cell cycle progression in Caulobacter crescentus.Cell Cycle. 2004 Oct;3(10):1252-4. doi: 10.4161/cc.3.10.1181. Epub 2004 Oct 17. Cell Cycle. 2004. PMID: 15467452 Review.
-
An intracellular compass spatially coordinates cell cycle modules in Caulobacter crescentus.Curr Opin Microbiol. 2016 Oct;33:131-139. doi: 10.1016/j.mib.2016.06.007. Epub 2016 Aug 10. Curr Opin Microbiol. 2016. PMID: 27517351 Free PMC article. Review.
Cited by
-
Bacterial polarity.Curr Opin Cell Biol. 2011 Feb;23(1):71-7. doi: 10.1016/j.ceb.2010.10.013. Epub 2010 Nov 20. Curr Opin Cell Biol. 2011. PMID: 21095111 Free PMC article.
-
Spatial perturbation with synthetic protein scaffold reveals robustness of asymmetric cell division.J Biomed Sci Eng. 2013 Feb;6(2):134-143. doi: 10.4236/jbise.2013.62017. J Biomed Sci Eng. 2013. PMID: 25750689 Free PMC article.
-
DNA Replication Initiation Is Blocked by a Distant Chromosome-Membrane Attachment.Curr Biol. 2015 Aug 17;25(16):2143-9. doi: 10.1016/j.cub.2015.06.058. Epub 2015 Aug 6. Curr Biol. 2015. PMID: 26255849 Free PMC article.
-
Synchronized Swarmers and Sticky Stalks: Caulobacter crescentus as a Model for Bacterial Cell Biology.J Bacteriol. 2023 Feb 22;205(2):e0038422. doi: 10.1128/jb.00384-22. Epub 2023 Jan 30. J Bacteriol. 2023. PMID: 36715542 Free PMC article. Review.
-
Cell cycle progression in Caulobacter requires a nucleoid-associated protein with high AT sequence recognition.Proc Natl Acad Sci U S A. 2016 Oct 4;113(40):E5952-E5961. doi: 10.1073/pnas.1612579113. Epub 2016 Sep 19. Proc Natl Acad Sci U S A. 2016. PMID: 27647925 Free PMC article.
References
-
- Aaron M, Charbon G, Lam H, Schwartz H, Vollmer W, Jacobs-Wagner C. 2007. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol Microbiol 64: 938–952. - PubMed
-
- Arias EE, Walter JC. 2007. Strength in numbers: Preventing rereplication via multiple mechanisms in eukaryotic cells. Genes Dev 21: 497–518. - PubMed
-
- Bell SP, Dutta A. 2002. DNA replication in eukaryotic cells. Annu Rev Biochem 71: 333–74. - PubMed
-
- Bellefontaine AF, Pierreux CE, Mertens P, Vandenhaute J, Letesson JJ, De Bolle X. 2002. Plasticity of a transcriptional regulation network among α-proteobacteria is supported by the identification of CtrA targets in Brucella abortus. Mol Microbiol 43: 945–960. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources