

Distinguishing direct versus indirect transcription factor-DNA interactions
- PMID: 19652015
- PMCID: PMC2775597
- DOI: 10.1101/gr.094144.109
Distinguishing direct versus indirect transcription factor-DNA interactions
Abstract
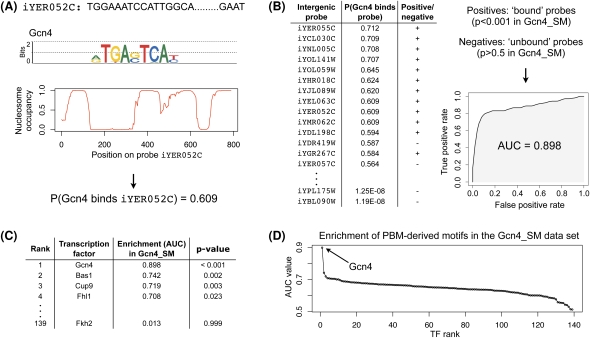
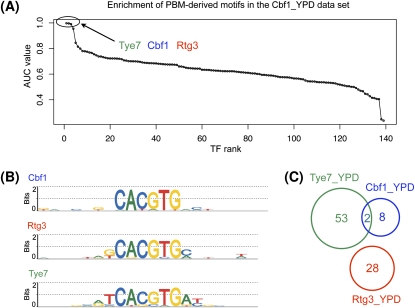
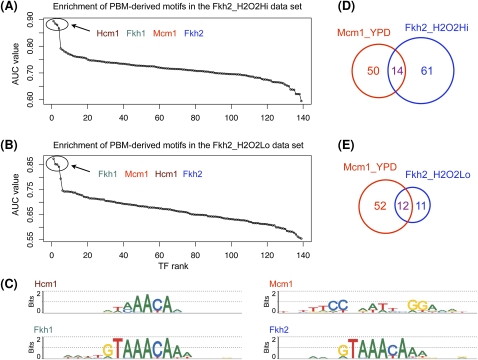
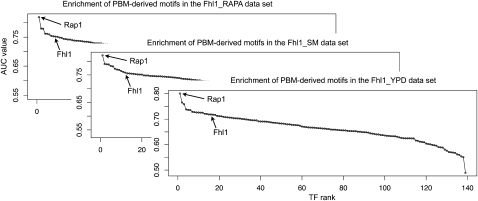
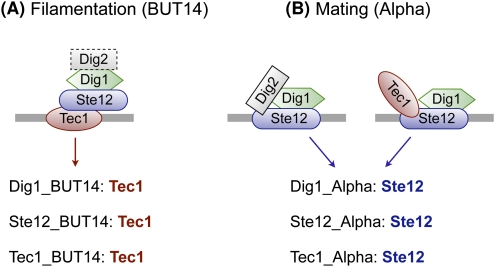
Transcriptional regulation is largely enacted by transcription factors (TFs) binding DNA. Large numbers of TF binding motifs have been revealed by ChIP-chip experiments followed by computational DNA motif discovery. However, the success of motif discovery algorithms has been limited when applied to sequences bound in vivo (such as those identified by ChIP-chip) because the observed TF-DNA interactions are not necessarily direct: Some TFs predominantly associate with DNA indirectly through protein partners, while others exhibit both direct and indirect binding. Here, we present the first method for distinguishing between direct and indirect TF-DNA interactions, integrating in vivo TF binding data, in vivo nucleosome occupancy data, and motifs from in vitro protein binding microarray experiments. When applied to yeast ChIP-chip data, our method reveals that only 48% of the data sets can be readily explained by direct binding of the profiled TF, while 16% can be explained by indirect DNA binding. In the remaining 36%, none of the motifs used in our analysis was able to explain the ChIP-chip data, either because the data were too noisy or because the set of motifs was incomplete. As more in vitro TF DNA binding motifs become available, our method could be used to build a complete catalog of direct and indirect TF-DNA interactions. Our method is not restricted to yeast or to ChIP-chip data, but can be applied in any system for which both in vivo binding data and in vitro DNA binding motifs are available.
Figures
Similar articles
-
High-resolution DNA-binding specificity analysis of yeast transcription factors.Genome Res. 2009 Apr;19(4):556-66. doi: 10.1101/gr.090233.108. Epub 2009 Jan 21. Genome Res. 2009. PMID: 19158363 Free PMC article.
-
Identifying cooperative transcription factors in yeast using multiple data sources.BMC Syst Biol. 2014;8 Suppl 5(Suppl 5):S2. doi: 10.1186/1752-0509-8-S5-S2. Epub 2014 Dec 12. BMC Syst Biol. 2014. PMID: 25559499 Free PMC article.
-
An algorithm for finding protein-DNA binding sites with applications to chromatin-immunoprecipitation microarray experiments.Nat Biotechnol. 2002 Aug;20(8):835-9. doi: 10.1038/nbt717. Epub 2002 Jul 8. Nat Biotechnol. 2002. PMID: 12101404
-
The different (sur)faces of Rap1p.Mol Genet Genomics. 2003 Mar;268(6):791-8. doi: 10.1007/s00438-002-0801-3. Epub 2003 Jan 25. Mol Genet Genomics. 2003. PMID: 12655405 Review.
-
A conserved role for transcription factor sumoylation in binding-site selection.Curr Genet. 2019 Dec;65(6):1307-1312. doi: 10.1007/s00294-019-00992-w. Epub 2019 May 15. Curr Genet. 2019. PMID: 31093693 Review.
Cited by
-
A structure-based Multiple-Instance Learning approach to predicting in vitro transcription factor-DNA interaction.BMC Genomics. 2015;16 Suppl 4(Suppl 4):S3. doi: 10.1186/1471-2164-16-S4-S3. Epub 2015 Apr 21. BMC Genomics. 2015. PMID: 25917392 Free PMC article.
-
Dry and wet approaches for genome-wide functional annotation of conventional and unconventional transcriptional activators.Comput Struct Biotechnol J. 2016 Jun 29;14:262-70. doi: 10.1016/j.csbj.2016.06.004. eCollection 2016. Comput Struct Biotechnol J. 2016. PMID: 27453771 Free PMC article. Review.
-
The orchestration of mammalian tissue morphogenesis through a series of coherent feed-forward loops.J Biol Chem. 2011 Dec 16;286(50):43259-71. doi: 10.1074/jbc.M111.264580. Epub 2011 Oct 13. J Biol Chem. 2011. PMID: 21998302 Free PMC article.
-
Computational approaches to understand transcription regulation in development.Biochem Soc Trans. 2023 Feb 27;51(1):1-12. doi: 10.1042/BST20210145. Biochem Soc Trans. 2023. PMID: 36695505 Free PMC article. Review.
-
Motif Enrichment Analysis: a unified framework and an evaluation on ChIP data.BMC Bioinformatics. 2010 Apr 1;11:165. doi: 10.1186/1471-2105-11-165. BMC Bioinformatics. 2010. PMID: 20356413 Free PMC article.
References
-
- Angermayr M, Oechsner U, Bandlow W. Reb1p-dependent DNA bending effects nucleosome positioning and constitutive transcription at the yeast profilin promoter. J Biol Chem. 2003;278:17918–17926. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous