

Fgf10 regulates transition period of cortical stem cell differentiation to radial glia controlling generation of neurons and basal progenitors
- PMID: 19607792
- PMCID: PMC2746711
- DOI: 10.1016/j.neuron.2009.06.006
Fgf10 regulates transition period of cortical stem cell differentiation to radial glia controlling generation of neurons and basal progenitors
Abstract
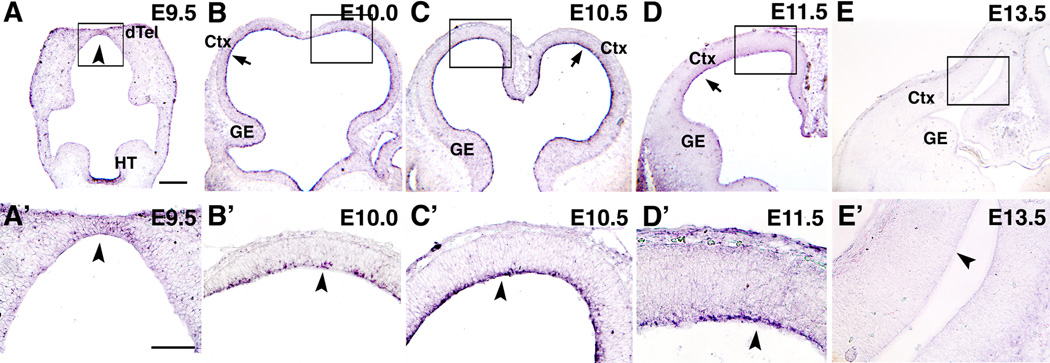
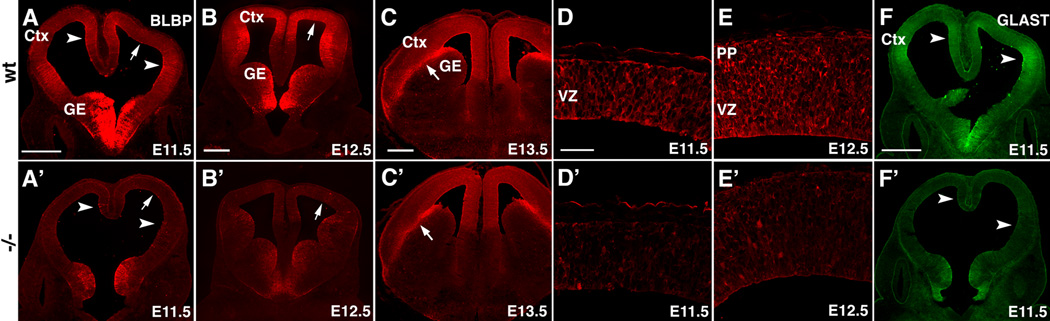
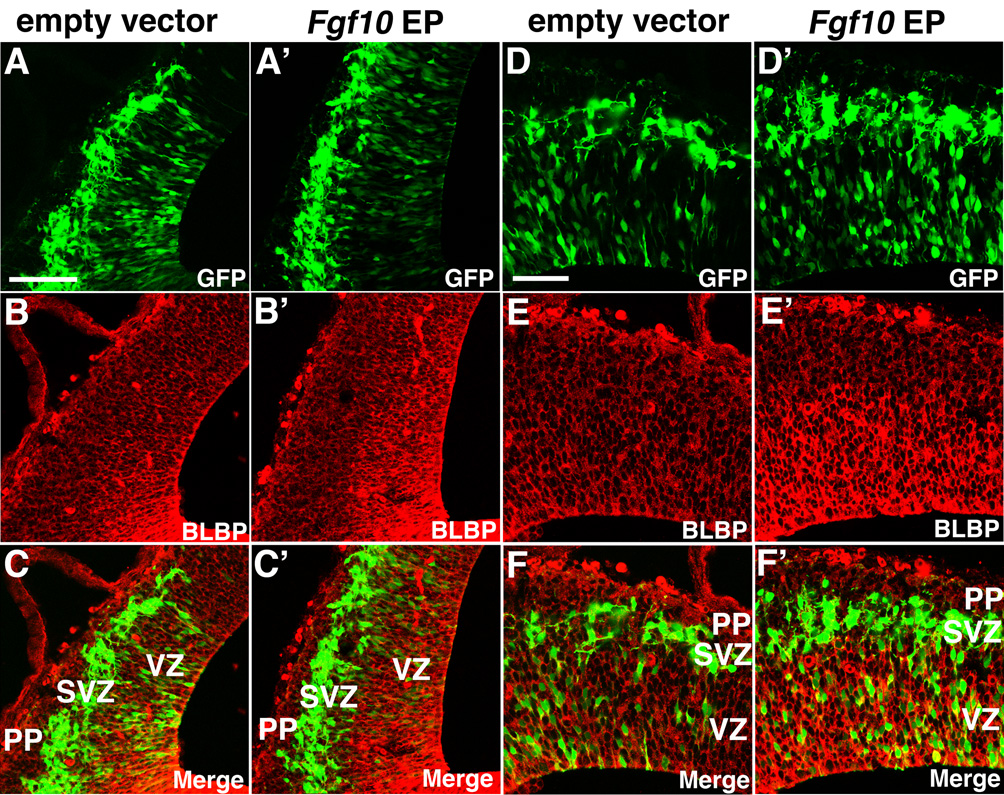
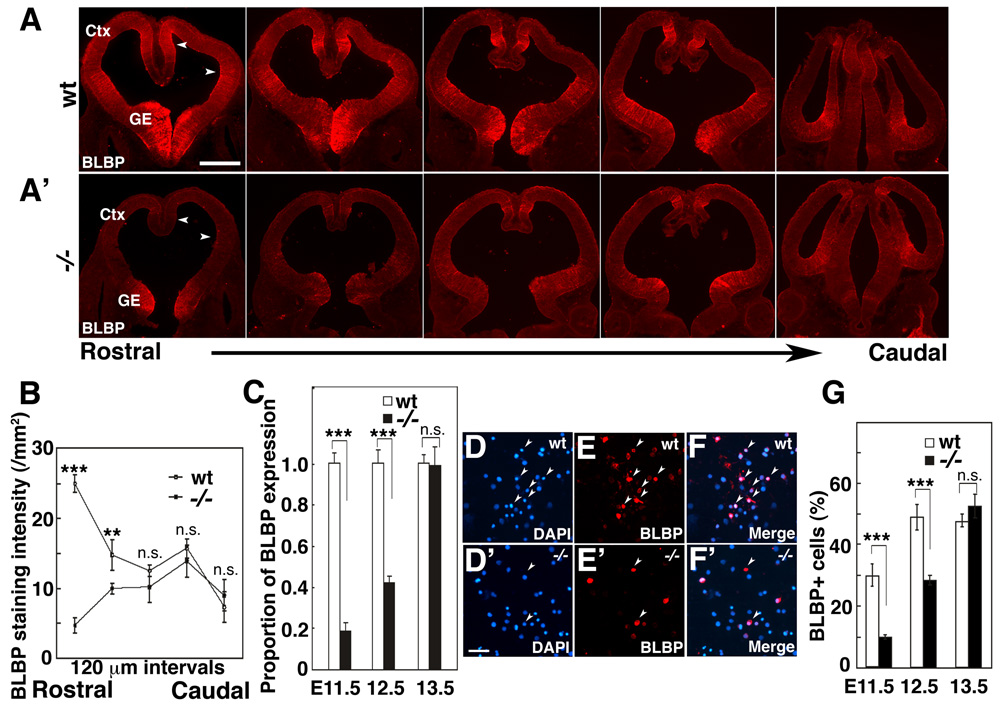
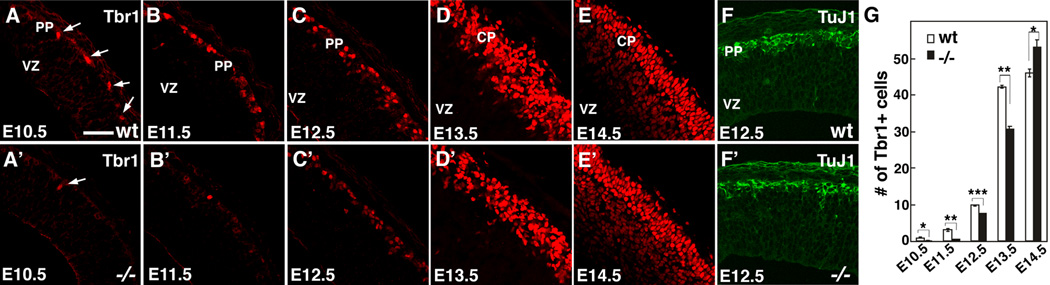
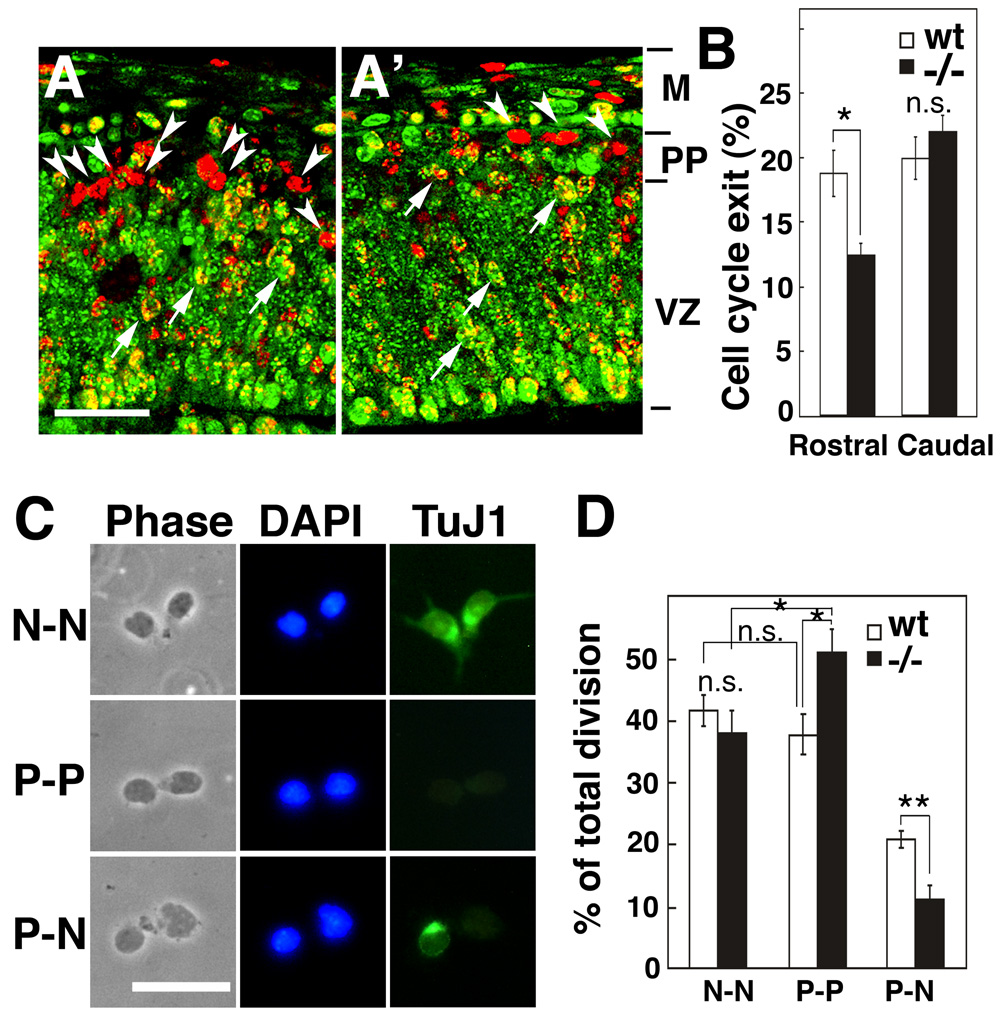
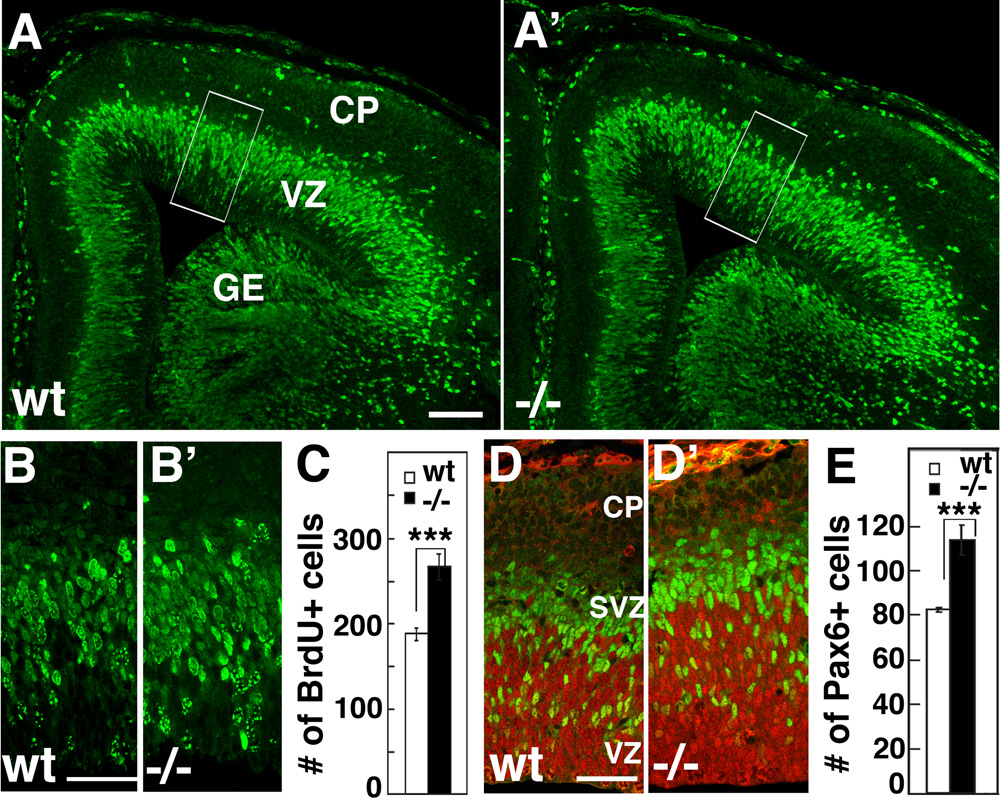
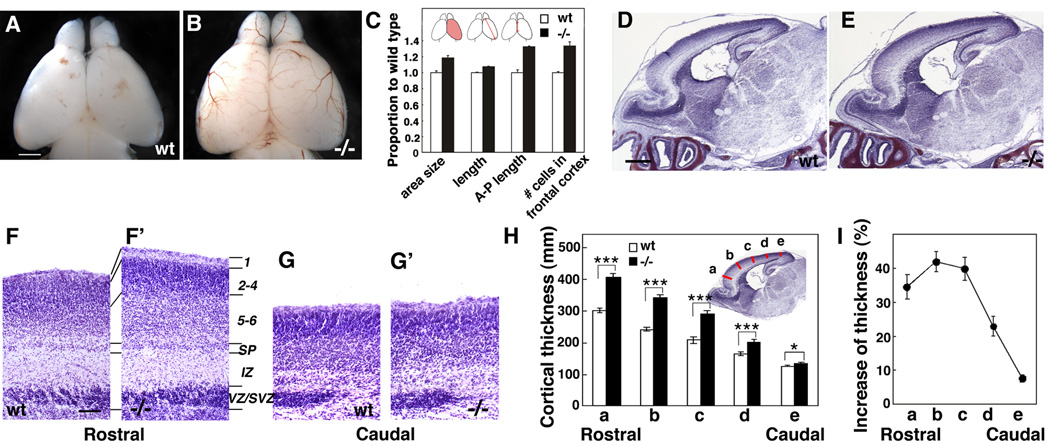
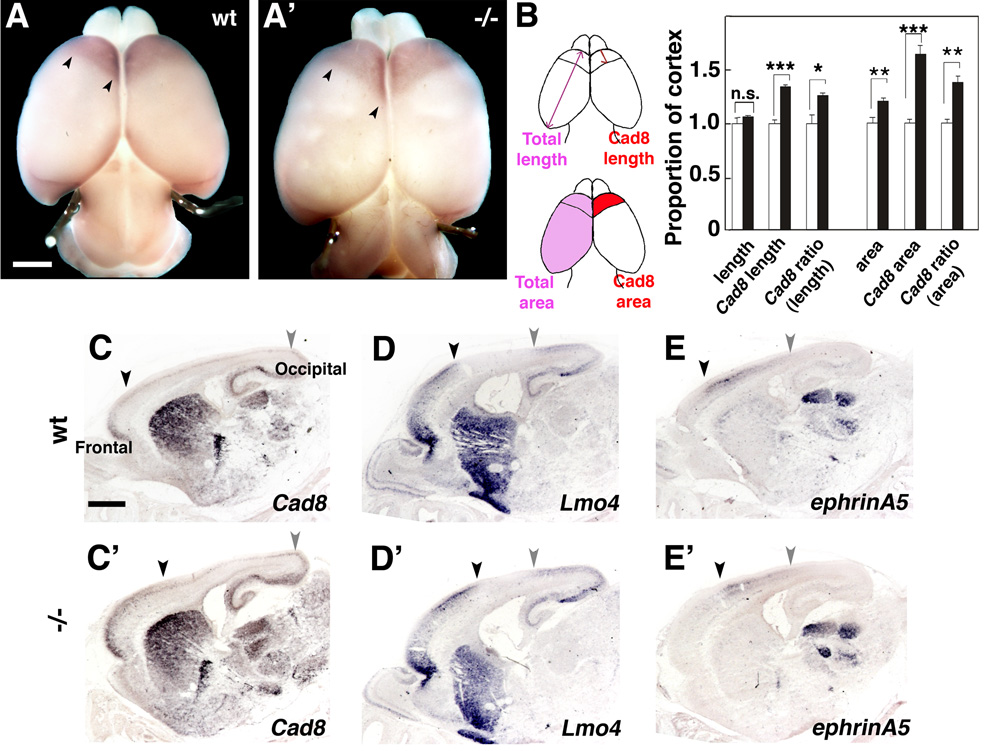
Radial glia (RG), the progenitors of cortical neurons and basal progenitors (BPs), differentiate from neuroepithelial cells (NCs) with stem cell properties. We show that the morphogen Fgf10 is transiently expressed by NCs coincident with the transition period of NC differentiation into RG. Targeted deletion of Fgf10 delays RG differentiation, whereas overexpression has opposing effects. Delayed RG differentiation in Fgf10 mutants occurs selectively in rostral cortex, paralleled by an extended period of symmetric NC divisions increasing progenitor number, coupled with delayed and initially diminished production of neurons and BPs. RG eventually differentiate in excess number and overproduce neurons and BPs rostrally resulting in tangential expansion of frontal areas and increased laminar thickness. Thus, transient Fgf10 expression regulates timely differentiation of RG, and through this function, determines both length of the early progenitor expansion phase and onset of neurogenesis and ultimately the number of progenitors and neurons fated to specific cortical areas.
Figures
Comment in
-
Baby got brain: Fgf10 sets rostral cortical size.Neuron. 2009 Jul 16;63(1):1-3. doi: 10.1016/j.neuron.2009.06.015. Neuron. 2009. PMID: 19607785
Similar articles
-
FGF signaling expands embryonic cortical surface area by regulating Notch-dependent neurogenesis.J Neurosci. 2011 Oct 26;31(43):15604-17. doi: 10.1523/JNEUROSCI.4439-11.2011. J Neurosci. 2011. PMID: 22031906 Free PMC article.
-
Selective cortical layering abnormalities and behavioral deficits in cortex-specific Pax6 knock-out mice.J Neurosci. 2009 Jul 1;29(26):8335-49. doi: 10.1523/JNEUROSCI.5669-08.2009. J Neurosci. 2009. PMID: 19571125 Free PMC article.
-
Tangentially migrating transient glutamatergic neurons control neurogenesis and maintenance of cerebral cortical progenitor pools.Cereb Cortex. 2012 Feb;22(2):403-16. doi: 10.1093/cercor/bhr122. Epub 2011 Jun 10. Cereb Cortex. 2012. PMID: 21666133 Free PMC article.
-
Neural stem and progenitor cells in cortical development.Novartis Found Symp. 2007;288:59-73; discussion 73-8, 96-8. Novartis Found Symp. 2007. PMID: 18494252 Review.
-
Role of intermediate progenitor cells in cerebral cortex development.Dev Neurosci. 2008;30(1-3):24-32. doi: 10.1159/000109848. Dev Neurosci. 2008. PMID: 18075251 Review.
Cited by
-
N-cadherin-based adherens junction regulates the maintenance, proliferation, and differentiation of neural progenitor cells during development.Cell Adh Migr. 2015;9(3):183-92. doi: 10.1080/19336918.2015.1005466. Epub 2015 Apr 14. Cell Adh Migr. 2015. PMID: 25869655 Free PMC article. Review.
-
Cortical gyrification induced by fibroblast growth factor 2 in the mouse brain.J Neurosci. 2013 Jun 26;33(26):10802-14. doi: 10.1523/JNEUROSCI.3621-12.2013. J Neurosci. 2013. PMID: 23804101 Free PMC article.
-
Coevolution of radial glial cells and the cerebral cortex.Glia. 2015 Aug;63(8):1303-19. doi: 10.1002/glia.22827. Epub 2015 Mar 23. Glia. 2015. PMID: 25808466 Free PMC article. Review.
-
Modeling Mammalian Commitment to the Neural Lineage Using Embryos and Embryonic Stem Cells.Front Physiol. 2019 Jul 11;10:705. doi: 10.3389/fphys.2019.00705. eCollection 2019. Front Physiol. 2019. PMID: 31354503 Free PMC article. Review.
-
Annual Research Review: Development of the cerebral cortex: implications for neurodevelopmental disorders.J Child Psychol Psychiatry. 2011 Apr;52(4):339-55. doi: 10.1111/j.1469-7610.2010.02307.x. Epub 2010 Aug 24. J Child Psychol Psychiatry. 2011. PMID: 20735793 Free PMC article. Review.
References
-
- Aaku-Saraste E, Hellwig A, Huttner WB. Loss of Occludin and functional tight junctions, but not ZO-1, during neural tube closure--remodeling of the neuroepithelium prior to neurogenesis. Dev Biol. 1996;180:664–679. - PubMed
-
- Anthony TE, Klein C, Fishell G, Heintz N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 2004;41:881–890. - PubMed
-
- Bachler M, Neubuser A. Expression of members of the Fgf family and their receptors during midfacial development. Mech Dev. 2001;100:313–316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
