

Functional unresponsiveness and replicative senescence of myeloid leukemia antigen-specific CD8+ T cells after allogeneic stem cell transplantation
- PMID: 19602548
- PMCID: PMC2722844
- DOI: 10.1158/1078-0432.CCR-08-3332
Functional unresponsiveness and replicative senescence of myeloid leukemia antigen-specific CD8+ T cells after allogeneic stem cell transplantation
Abstract
Purpose: The therapeutic effect of allogeneic hematopoietic stem cell transplantation (HSCT) for patients with myeloid malignancies has been attributed in part to a graft-versus-leukemia effect that is dependent on donor T lymphocytes. CD8(+) T-cell responses to MHC class I-restricted tumor epitopes, not just allogeneic antigens, may help mediate antileukemia effects after HSCT, but the specificity and function of such cells are not completely understood.
Experimental design: We examined the diversity, phenotype, and functional potential of leukemia-associated antigen-specific CD8(+) T cells in patients with myeloid leukemia following allogeneic HSCT. Screening for antigen-specific T cells was accomplished with a peptide/MHC tetramer library.
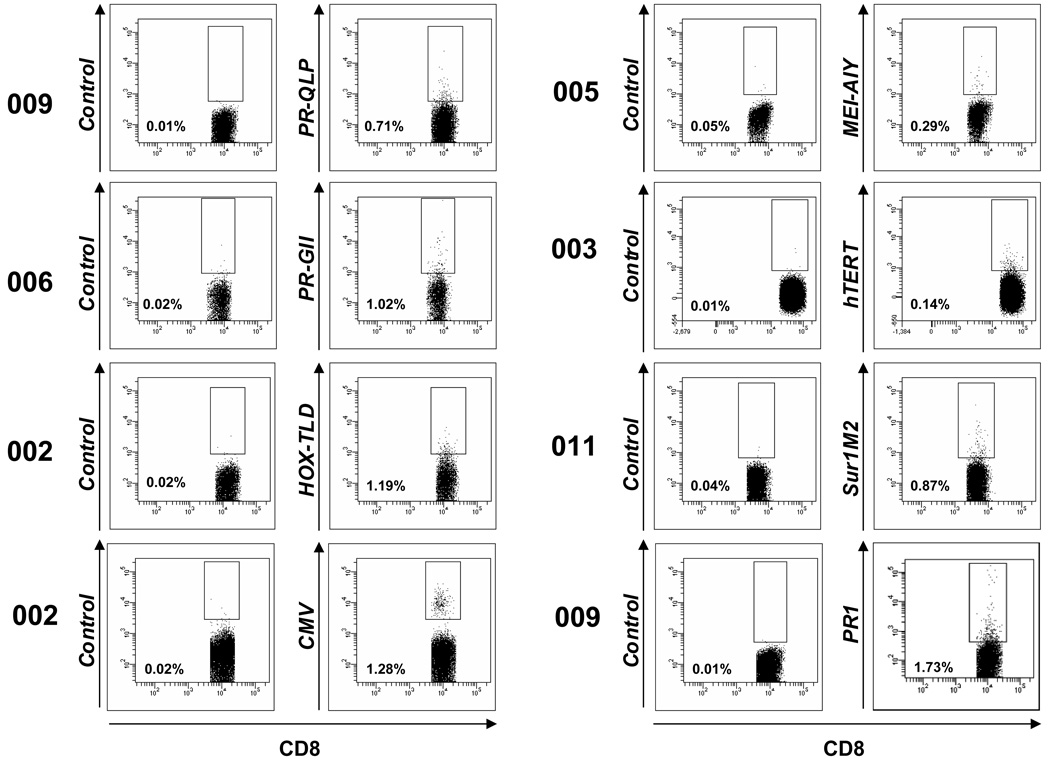
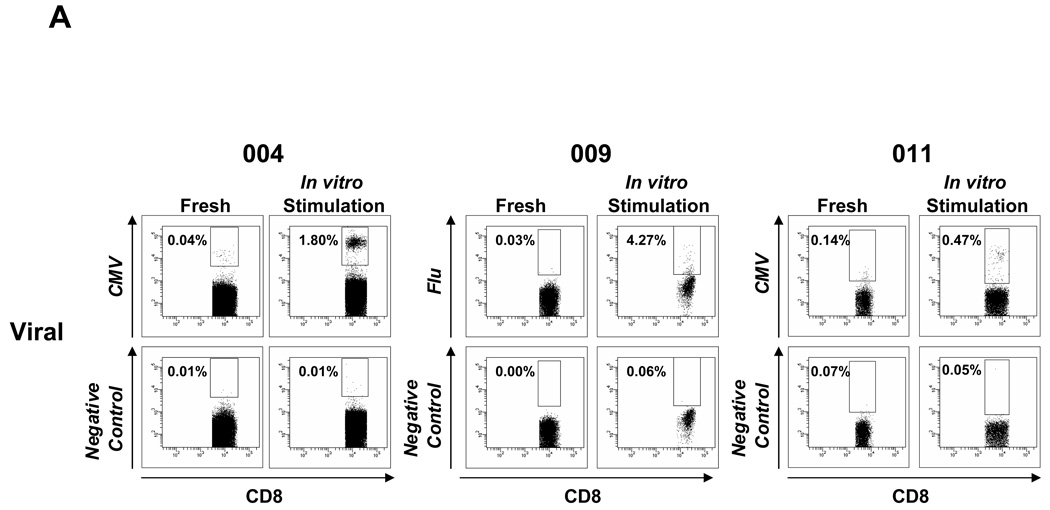
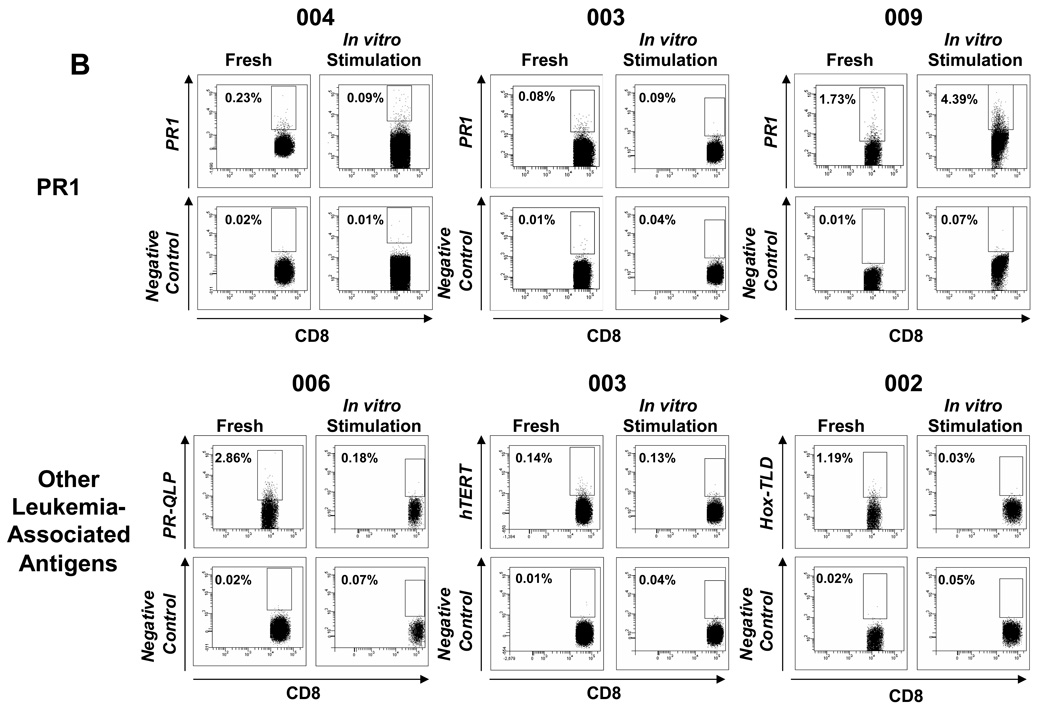
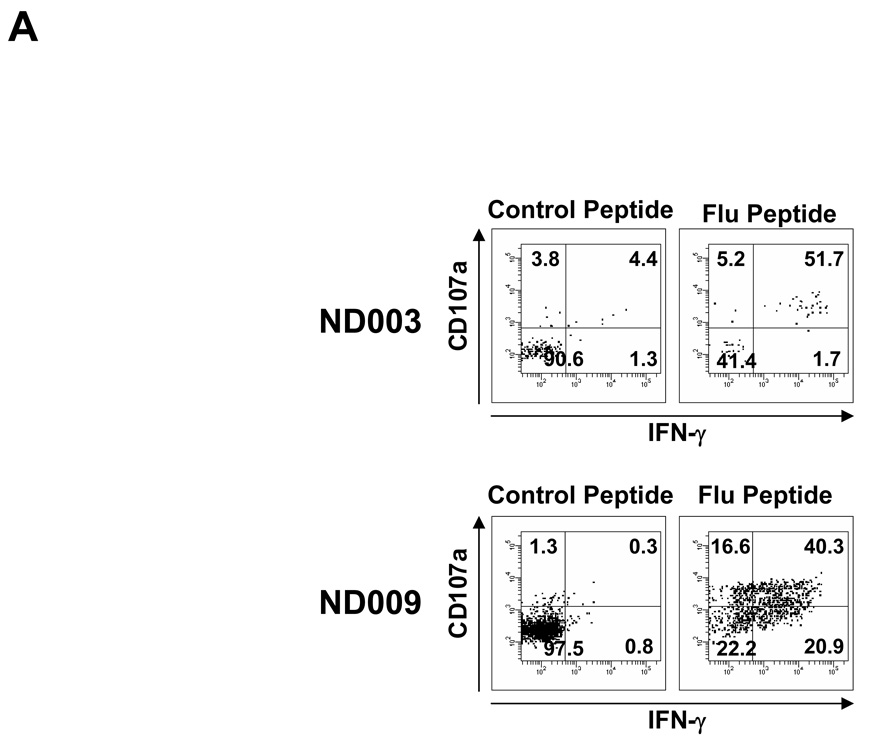
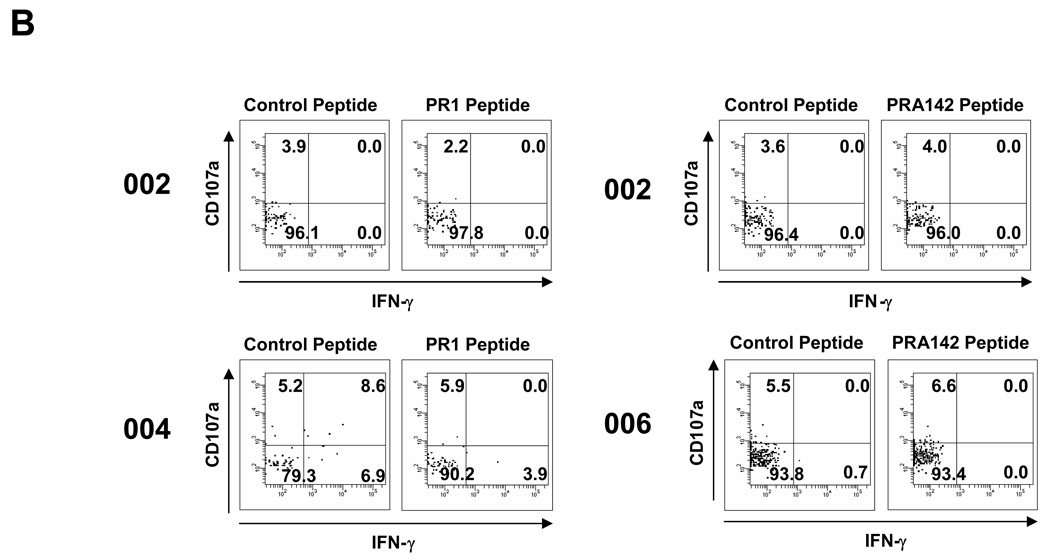
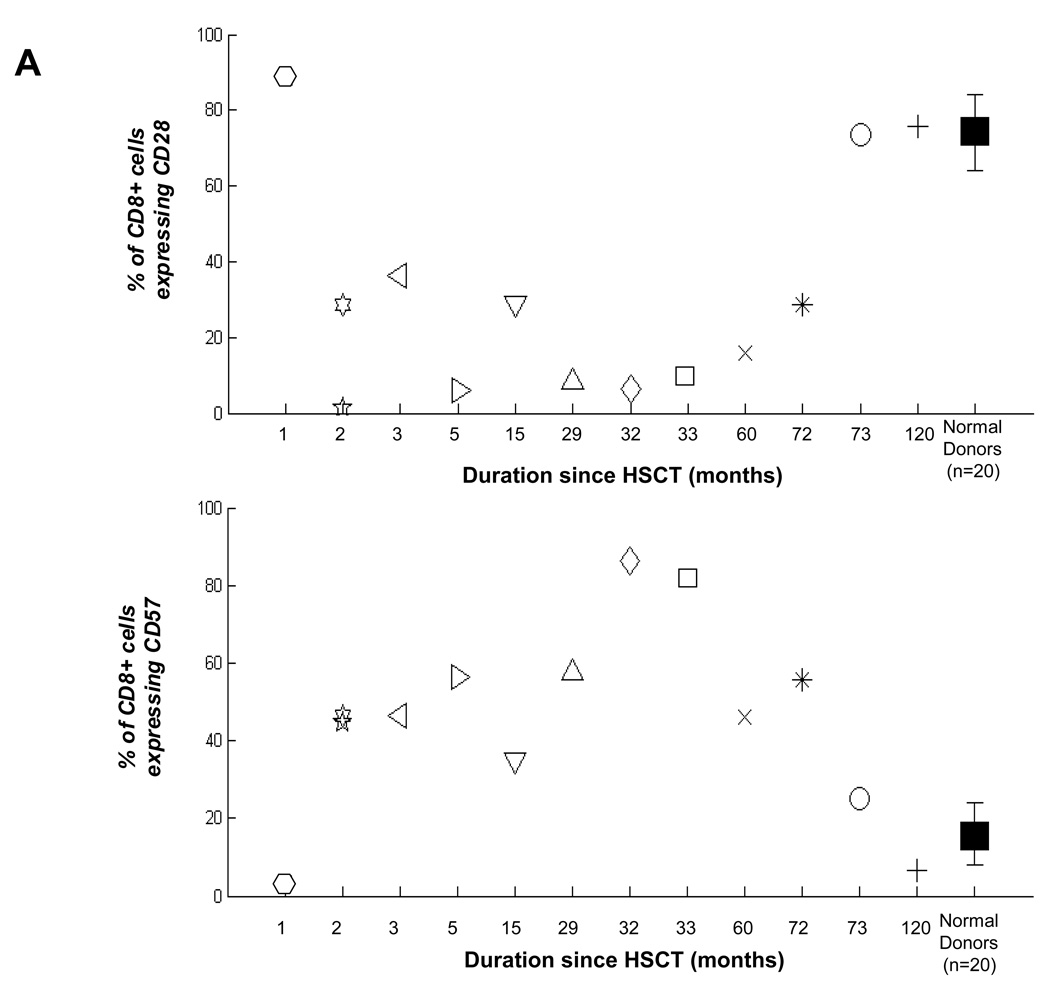
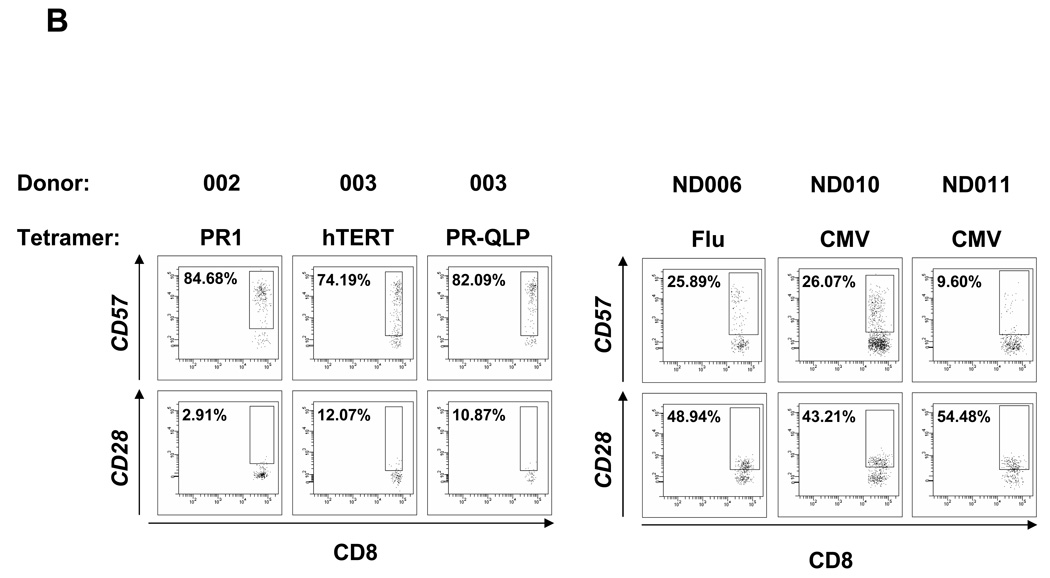
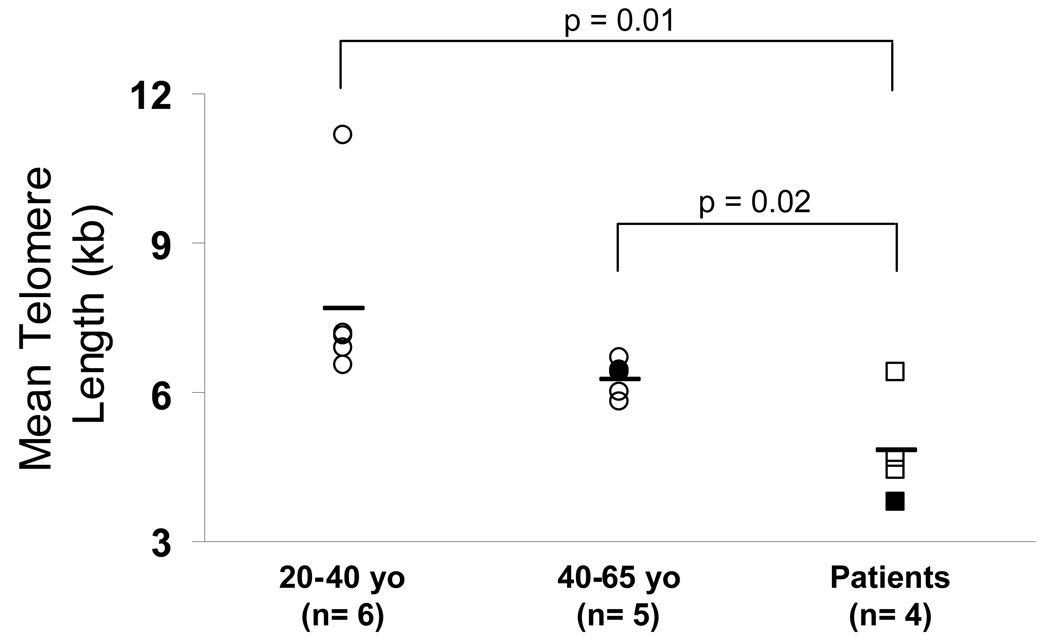
Results: Patients with acute myelogenous leukemia or chronic myelogenous leukemia in remission following HSCT exhibited significant numbers of peripheral blood CD8(+) T cells that recognized varying combinations of epitopes derived from leukemia-associated antigens. However, these cells failed to proliferate, release cytokines, or degranulate in response to antigen-specific stimuli. As early as 2 months after HSCT, CD8(+) T cells from patients were predominantly CD28(-) CD57(+) and had relatively short telomeres, consistent with cellular senescence.
Conclusions: Circulating leukemia-specific CD8(+) T cells are prominent in myeloid leukemia patients after HSCT, but such cells are largely functionally unresponsive, most likely due to replicative senescence. These findings carry important implications for the understanding of the graft-versus-leukemia effect and for the rational design of immunotherapeutic strategies for patients with myeloid leukemias.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Similar articles
-
A frameshift polymorphism in P2X5 elicits an allogeneic cytotoxic T lymphocyte response associated with remission of chronic myeloid leukemia.J Clin Invest. 2005 Dec;115(12):3506-16. doi: 10.1172/JCI24832. J Clin Invest. 2005. PMID: 16322791 Free PMC article.
-
CD8(+) T-cell immunity against cancer-testis antigens develops following allogeneic stem cell transplantation and reveals a potential mechanism for the graft-versus-leukemia effect.Haematologica. 2010 Sep;95(9):1572-8. doi: 10.3324/haematol.2009.019539. Epub 2010 May 11. Haematologica. 2010. PMID: 20460638 Free PMC article.
-
Efficient activation of LRH-1-specific CD8+ T-cell responses from transplanted leukemia patients by stimulation with P2X5 mRNA-electroporated dendritic cells.J Immunother. 2009 Jul-Aug;32(6):539-51. doi: 10.1097/CJI.0b013e3181987c22. J Immunother. 2009. PMID: 19483655
-
Use of chimeric antigen receptor T cells in allogeneic hematopoietic stem cell transplantation.Immunotherapy. 2019 Jan;11(1):37-44. doi: 10.2217/imt-2018-0089. Immunotherapy. 2019. PMID: 30702011 Review.
-
A Case of Multiple Sclerosis-Like Relapsing Remitting Encephalomyelitis Following Allogeneic Hematopoietic Stem Cell Transplantation and a Review of the Published Literature.Front Immunol. 2020 May 5;11:668. doi: 10.3389/fimmu.2020.00668. eCollection 2020. Front Immunol. 2020. PMID: 32431694 Free PMC article. Review.
Cited by
-
Anti-tumor immunity: myeloid leukocytes control the immune landscape.Cell Immunol. 2012 Jul-Aug;278(1-2):21-6. doi: 10.1016/j.cellimm.2012.06.014. Epub 2012 Jul 14. Cell Immunol. 2012. PMID: 23121972 Free PMC article. Review.
-
Precision in donor selection: Identifying ideal stem-cell donors through their T cells.Exp Hematol. 2016 Nov;44(11):1020-1023. doi: 10.1016/j.exphem.2016.07.013. Epub 2016 Aug 2. Exp Hematol. 2016. PMID: 27496363 Free PMC article. Review.
-
Novel Therapies for Acute Myeloid Leukemia: Are We Finally Breaking the Deadlock?Target Oncol. 2017 Aug;12(4):413-447. doi: 10.1007/s11523-017-0503-8. Target Oncol. 2017. PMID: 28664386 Review.
-
Delivery of progenitors to the thymus limits T-lineage reconstitution after bone marrow transplantation.Blood. 2011 Aug 18;118(7):1962-70. doi: 10.1182/blood-2010-12-324954. Epub 2011 Jun 9. Blood. 2011. PMID: 21659540 Free PMC article.
-
Characterization of immunologic properties of a second HLA-A2 epitope from a granule protease in CML patients and HLA-A2 transgenic mice.Blood. 2011 Aug 25;118(8):2159-69. doi: 10.1182/blood-2011-04-349951. Epub 2011 Jun 30. Blood. 2011. PMID: 21719601 Free PMC article. Clinical Trial.
References
-
- Wu CJ, Ritz J. Induction of tumor immunity following allogeneic stem cell transplantation. Adv Immunol. 2006;90:133–173. - PubMed
-
- Molldrem JJ, Lee PP, Wang C, Felio K, Kantarjian HM, Champlin RE, Davis MM. Evidence that specific T lymphocytes may participate in the elimination of chronic myelogenous leukemia. Nat Med. 2000;6:1018–1023. - PubMed
-
- Rusakiewicz S, Molldrem JJ. Immunotherapeutic peptide vaccination with leukemia-associated antigens. Curr Opin Immunol. 2006;18:599–604. - PubMed
-
- Molldrem J, Dermime S, Parker K, Jiang YZ, Mavroudis D, Hensel N, Fukushima P, Barrett AJ. Targeted T-cell therapy for human leukemia: cytotoxic T lymphocytes specific for a peptide derived from proteinase 3 preferentially lyse human myeloid leukemia cells. Blood. 1996;88:2450–2457. - PubMed
-
- Gannage M, Abel M, Michallet AS, Delluc S, Lambert M, Giraudier S, Kratzer R, Niedermann G, Saveanu L, Guilhot F, Camoin L, Varet B, Buzyn A, Caillat-Zucman S. Ex vivo characterization of multiepitopic tumor-specific CD8 T cells in patients with chronic myeloid leukemia: implications for vaccine development and adoptive cellular immunotherapy. J Immunol. 2005;174:8210–8218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
