

Conformational changes associated with template commitment in ATP-dependent chromatin remodeling by ISW2
- PMID: 19595716
- PMCID: PMC2721901
- DOI: 10.1016/j.molcel.2009.05.013
Conformational changes associated with template commitment in ATP-dependent chromatin remodeling by ISW2
Abstract
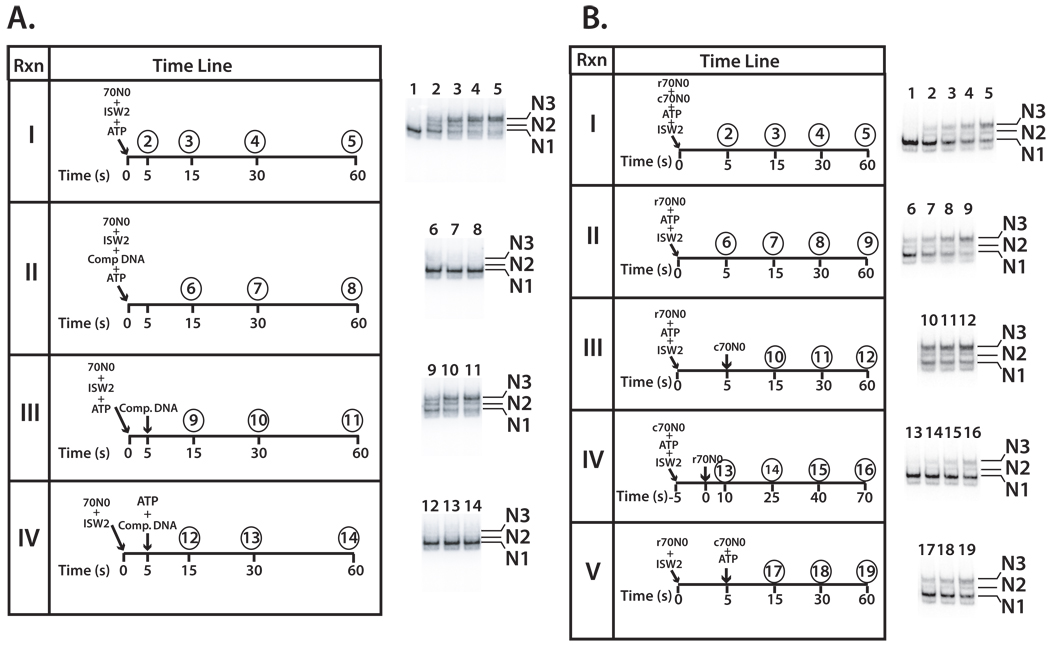
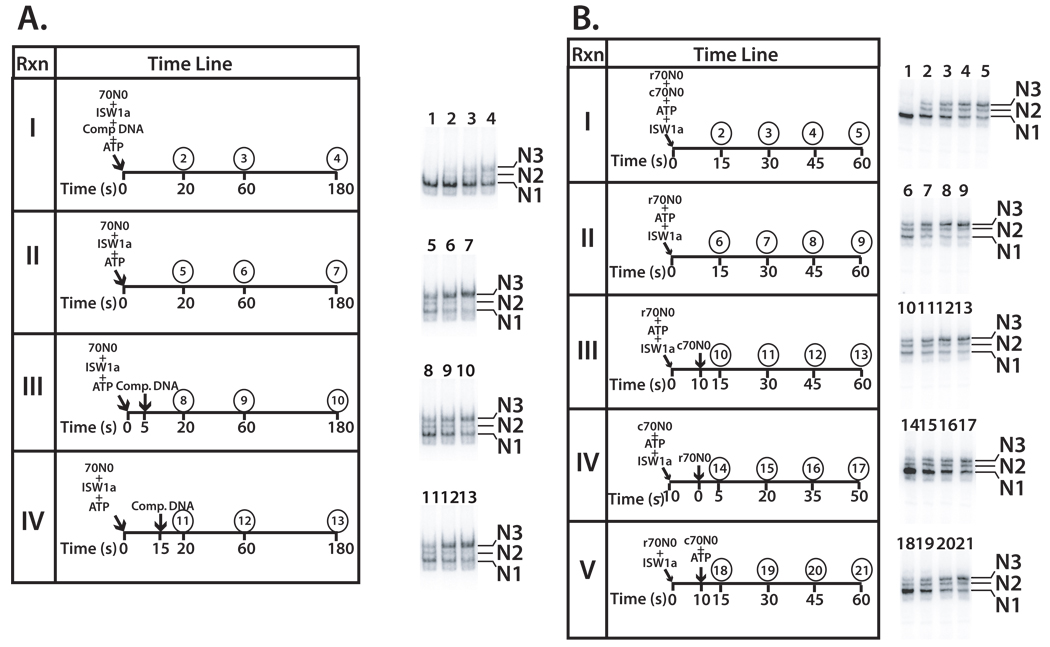
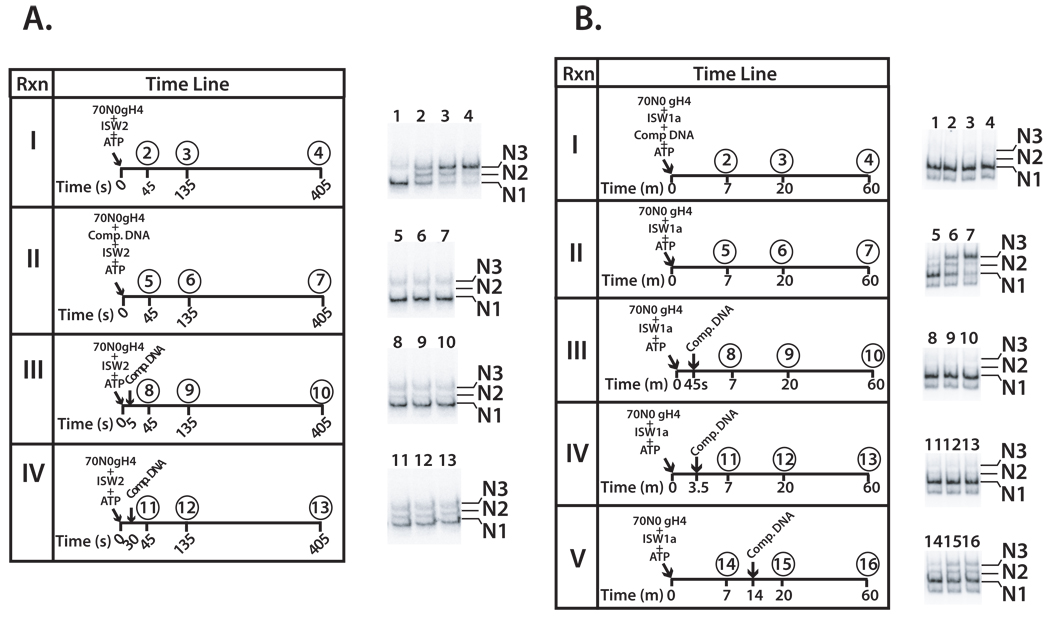
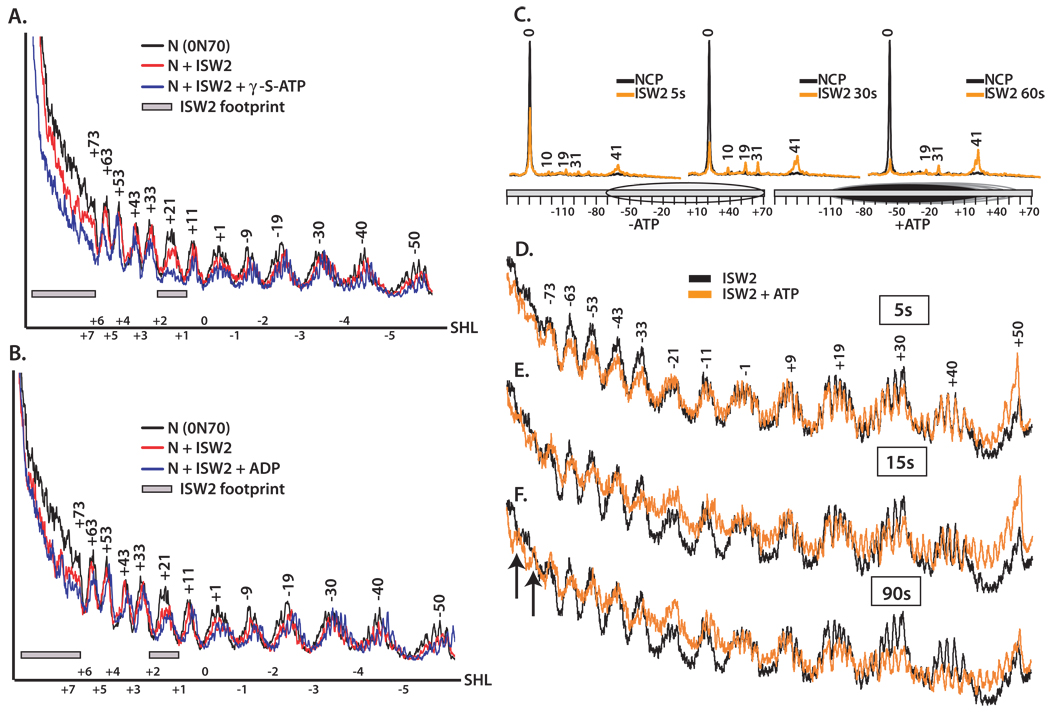
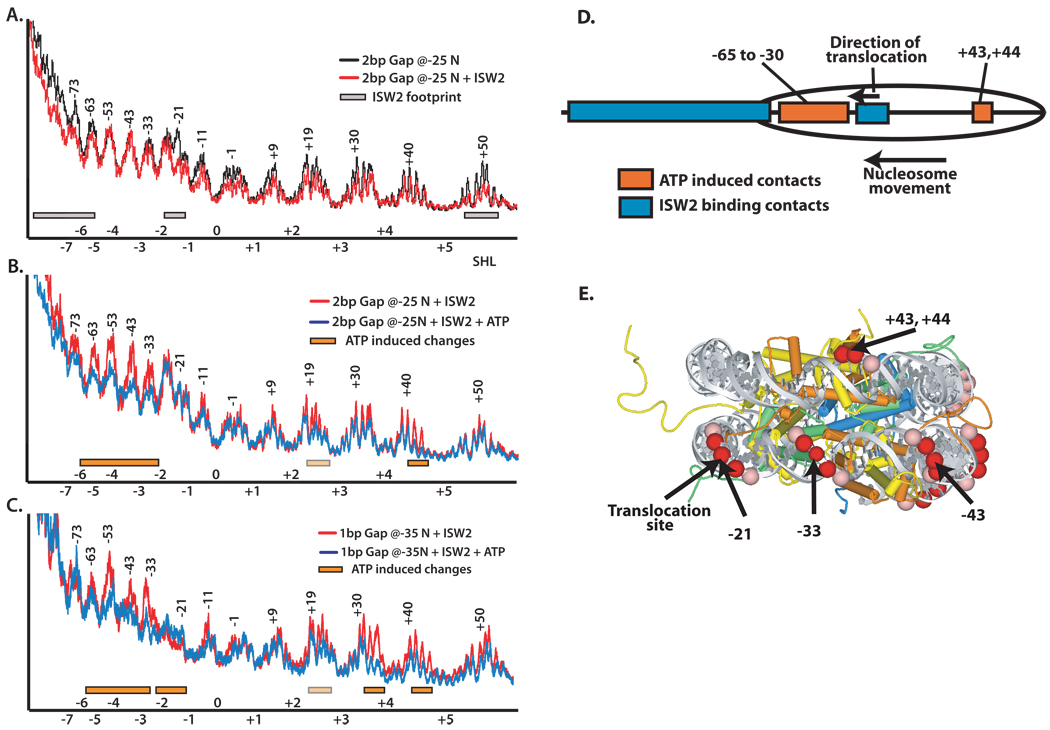
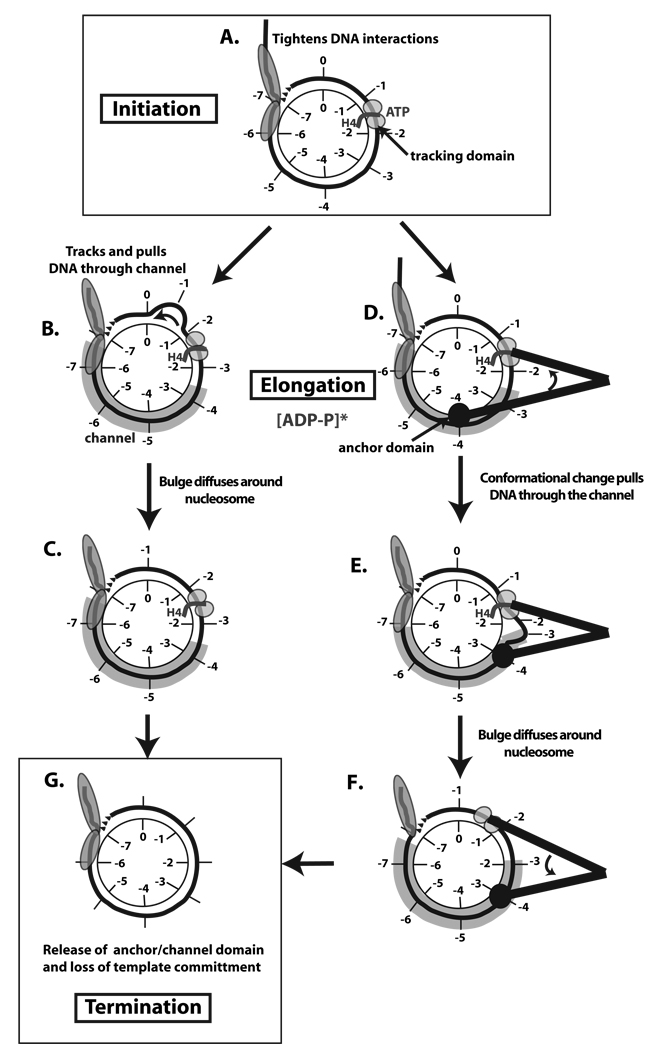
Distinct stages in ATP-dependent chromatin remodeling are found as ISW2, an ISWI-type complex, forms a stable and processive complex with nucleosomes upon hydrolysis of ATP. There are two conformational changes of the ISW2-nucleosome complex associated with binding and hydrolysis of ATP. The initial binding of ISW2 to extranucleosomal DNA, to the entry site, and near the dyad axis of the nucleosome is enhanced by ATP binding, whereas subsequent ATP hydrolysis is required for template commitment and causes ISW2 to expand its interactions with nucleosomal DNA to an entire gyre of the nucleosome and a short approximately 3-4 bp site on the other gyre. The histone-fold-like subunit Dpb4 associates with nucleosomal DNA approximately 15 bp from the ATPase domain as part of this change and may help to disrupt histone-DNA interactions. These additional contacts are independent of the ATPase domain tracking along nucleosomal DNA and are maintained as ISW2 moves nucleosomes on DNA.
Figures

Similar articles
-
A proposal for kinetic proof reading by ISWI family chromatin remodeling motors.Curr Opin Chem Biol. 2010 Oct;14(5):660-5. doi: 10.1016/j.cbpa.2010.08.001. Epub 2010 Sep 15. Curr Opin Chem Biol. 2010. PMID: 20833099 Free PMC article. Review.
-
Chromatin remodeling by ISW2 and SWI/SNF requires DNA translocation inside the nucleosome.Nat Struct Mol Biol. 2006 Apr;13(4):339-46. doi: 10.1038/nsmb1071. Epub 2006 Mar 5. Nat Struct Mol Biol. 2006. PMID: 16518397
-
The Dpb4 subunit of ISW2 is anchored to extranucleosomal DNA.J Biol Chem. 2007 Jul 6;282(27):19418-25. doi: 10.1074/jbc.M700640200. Epub 2007 May 9. J Biol Chem. 2007. PMID: 17491017
-
Histone Octamer Structure Is Altered Early in ISW2 ATP-Dependent Nucleosome Remodeling.Cell Rep. 2019 Jul 2;28(1):282-294.e6. doi: 10.1016/j.celrep.2019.05.106. Cell Rep. 2019. PMID: 31269447
-
Snf2-family proteins: chromatin remodellers for any occasion.Curr Opin Chem Biol. 2011 Oct;15(5):649-56. doi: 10.1016/j.cbpa.2011.07.022. Epub 2011 Aug 20. Curr Opin Chem Biol. 2011. PMID: 21862382 Free PMC article. Review.
Cited by
-
A proposal for kinetic proof reading by ISWI family chromatin remodeling motors.Curr Opin Chem Biol. 2010 Oct;14(5):660-5. doi: 10.1016/j.cbpa.2010.08.001. Epub 2010 Sep 15. Curr Opin Chem Biol. 2010. PMID: 20833099 Free PMC article. Review.
-
The chromatin remodeller ACF acts as a dimeric motor to space nucleosomes.Nature. 2009 Dec 24;462(7276):1016-21. doi: 10.1038/nature08621. Nature. 2009. PMID: 20033039 Free PMC article.
-
Mechanisms for ATP-dependent chromatin remodelling: the means to the end.FEBS J. 2011 Oct;278(19):3579-95. doi: 10.1111/j.1742-4658.2011.08281.x. Epub 2011 Sep 8. FEBS J. 2011. PMID: 21810178 Free PMC article. Review.
-
Quantitative determination of binding of ISWI to nucleosomes and DNA shows allosteric regulation of DNA binding by nucleotides.Biochemistry. 2014 Jul 15;53(27):4334-45. doi: 10.1021/bi500224t. Epub 2014 Jun 30. Biochemistry. 2014. PMID: 24898734 Free PMC article.
-
Human ISWI chromatin-remodeling complexes sample nucleosomes via transient binding reactions and become immobilized at active sites.Proc Natl Acad Sci U S A. 2010 Nov 16;107(46):19873-8. doi: 10.1073/pnas.1003438107. Epub 2010 Oct 25. Proc Natl Acad Sci U S A. 2010. PMID: 20974961 Free PMC article.
References
-
- Cramer P, Bushnell DA, Fu J, Gnatt AL, Maier-Davis B, Thompson NE, Burgess RR, Edwards AM, David PR, Kornberg RD. Architecture of RNA polymerase II and implications for the transcription mechanism. Science. 2000;288:640–649. - PubMed
-
- Cramer P, Bushnell DA, Kornberg RD. Structural basis of transcription: RNA polymerase II at 2.8 angstrom resolution. Science. 2001;292:1863–1876. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
