

Divergent S phase checkpoint activation arising from prereplicative complex deficiency controls cell survival
- PMID: 19587119
- PMCID: PMC2735493
- DOI: 10.1091/mbc.e09-01-0022
Divergent S phase checkpoint activation arising from prereplicative complex deficiency controls cell survival
Abstract
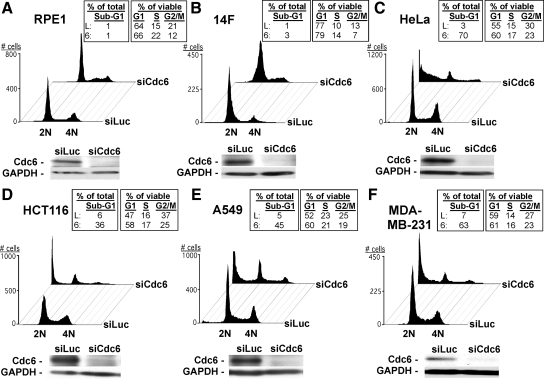
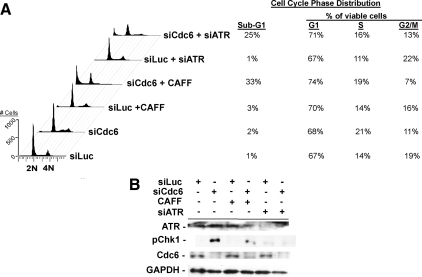
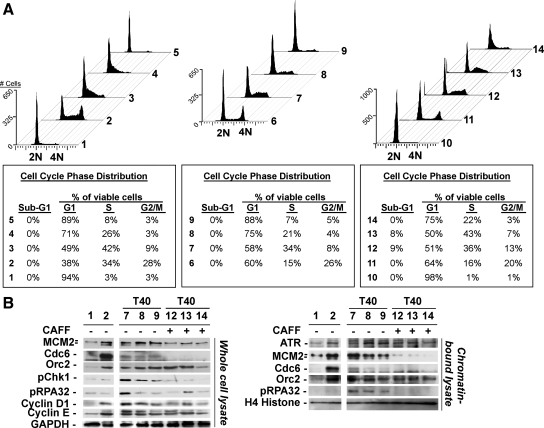
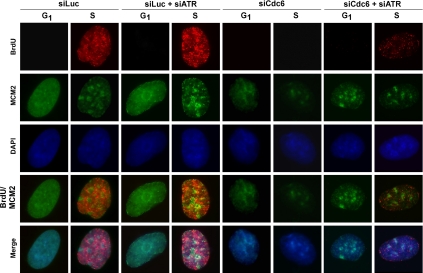
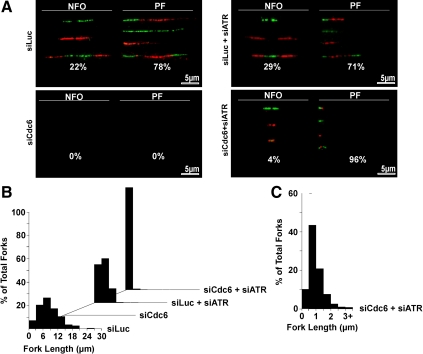
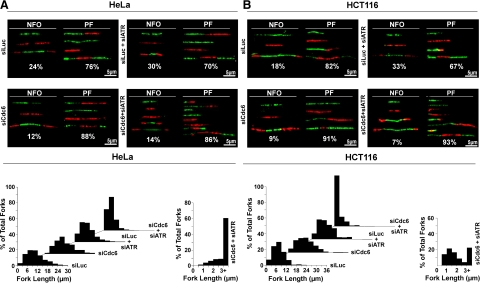
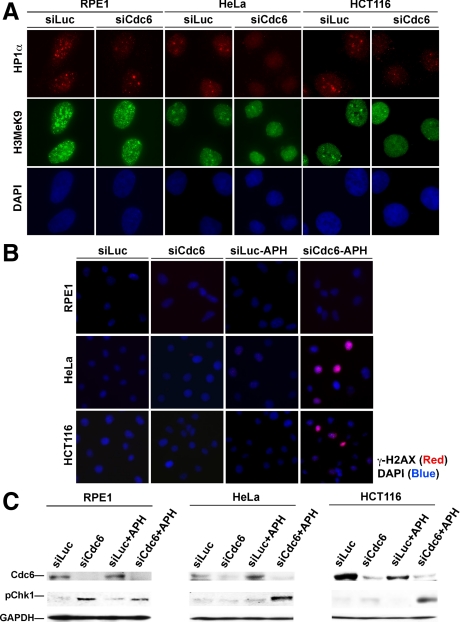
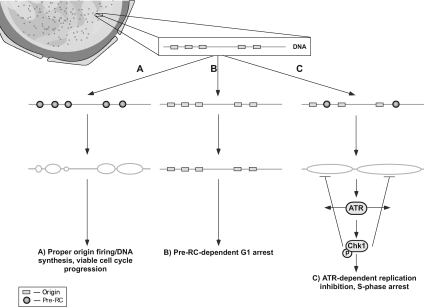
The DNA replication machinery plays additional roles in S phase checkpoint control, although the identities of the replication proteins involved in checkpoint activation remain elusive. Here, we report that depletion of the prereplicative complex (pre-RC) protein Cdc6 causes human nontransformed diploid cells to arrest nonlethally in G1-G1/S and S phase, whereas multiple cancer cell lines undergo G1-G1/S arrest and cell death. These divergent phenotypes are dependent on the activation, or lack thereof, of an ataxia telangiectasia and Rad3-related (ATR)-dependent S phase checkpoint that inhibits replication fork progression. Although pre-RC deficiency induces chromatin structural alterations in both nontransformed and cancer cells that normally lead to ATR checkpoint activation, the sensor mechanisms in cancer cells seem to be compromised such that higher levels of DNA replication stress/damage are required to trigger checkpoint response. Our results suggest that therapy-induced disruption of pre-RC function might exert selective cytotoxic effects on tumor cells in human patients.
Figures
Similar articles
-
The ATR-mediated S phase checkpoint prevents rereplication in mammalian cells when licensing control is disrupted.J Cell Biol. 2007 Nov 19;179(4):643-57. doi: 10.1083/jcb.200704138. J Cell Biol. 2007. PMID: 18025301 Free PMC article.
-
CDC6 interaction with ATR regulates activation of a replication checkpoint in higher eukaryotic cells.J Cell Sci. 2010 Jan 15;123(Pt 2):225-35. doi: 10.1242/jcs.058693. J Cell Sci. 2010. PMID: 20048340
-
An ATR- and Chk1-dependent S checkpoint inhibits replicon initiation following UVC-induced DNA damage.Mol Cell Biol. 2002 Dec;22(24):8552-61. doi: 10.1128/MCB.22.24.8552-8561.2002. Mol Cell Biol. 2002. PMID: 12446774 Free PMC article.
-
Cyclin-dependent kinases and S phase control in mammalian cells.Cell Cycle. 2003 Jul-Aug;2(4):316-24. Cell Cycle. 2003. PMID: 12851482 Review.
-
Evidence that the ATR/Chk1 pathway maintains normal replication fork progression during unperturbed S phase.Cell Cycle. 2006 Oct;5(19):2203-9. doi: 10.4161/cc.5.19.3256. Epub 2006 Oct 1. Cell Cycle. 2006. PMID: 16969104 Review.
Cited by
-
ImmunoCellCycle-ID: A high-precision immunofluorescence-based method for cell cycle identification.bioRxiv [Preprint]. 2024 Aug 15:2024.08.14.607961. doi: 10.1101/2024.08.14.607961. bioRxiv. 2024. Update in: J Cell Sci. 2024 Nov 15;137(22):jcs263414. doi: 10.1242/jcs.263414 PMID: 39185179 Free PMC article. Updated. Preprint.
-
Mechanism of cancer cell death induced by depletion of an essential replication regulator.PLoS One. 2012;7(5):e36372. doi: 10.1371/journal.pone.0036372. Epub 2012 May 4. PLoS One. 2012. PMID: 22574151 Free PMC article.
-
Mitotic UV irradiation induces a DNA replication-licensing defect that potentiates G1 arrest response.PLoS One. 2015 Mar 23;10(3):e0120553. doi: 10.1371/journal.pone.0120553. eCollection 2015. PLoS One. 2015. PMID: 25798850 Free PMC article.
-
Reversible and effective cell cycle synchronization method for studying stage-specific investigations.bioRxiv [Preprint]. 2024 Sep 3:2024.09.02.610832. doi: 10.1101/2024.09.02.610832. bioRxiv. 2024. PMID: 39282459 Free PMC article. Preprint.
-
Suppression of Reserve MCM Complexes Chemosensitizes to Gemcitabine and 5-Fluorouracil.Mol Cancer Res. 2015 Sep;13(9):1296-305. doi: 10.1158/1541-7786.MCR-14-0464. Epub 2015 Jun 10. Mol Cancer Res. 2015. PMID: 26063742 Free PMC article.
References
-
- Abraham R. T. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev. 2001;15:2177–2196. - PubMed
-
- Anantha R. W., Vassin V. M., Borowiec J. A. Sequential and synergistic modification of human RPA stimulates chromosomal DNA repair. J. Biol. Chem. 2007;282:35910–35923. - PubMed
-
- Ayoub N., Jeyasekharan A. D., Bernal J. A., Venkitaraman A. R. HP1-beta mobilization promotes chromatin changes that initiate the DNA damage response. Nature. 2008;453:682–686. - PubMed
-
- Bartek J., Bartkova J., Lukas J. DNA damage signalling guards against activated oncogenes and tumour progression. Oncogene. 2007a;26:7773–7779. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
