

Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant
- PMID: 19564349
- PMCID: PMC2715098
- DOI: 10.1084/jem.20090247
Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant
Abstract
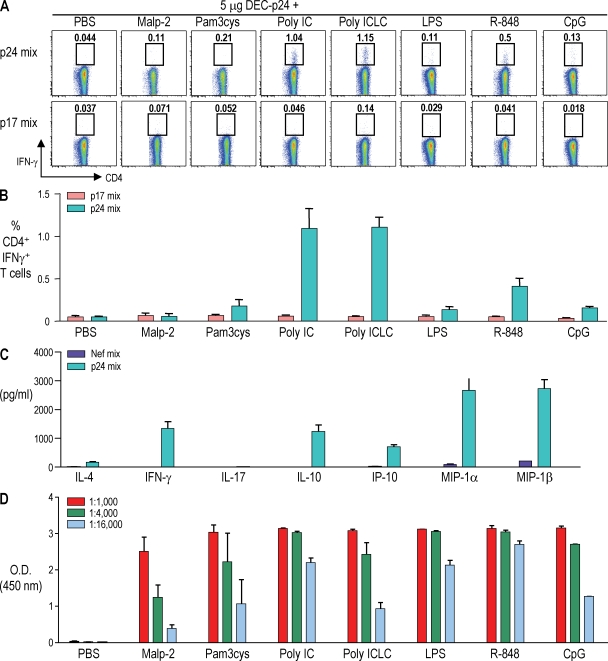
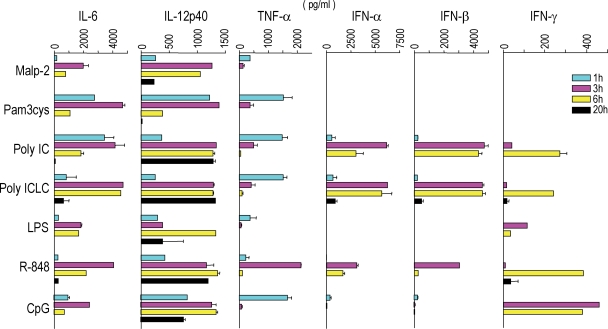
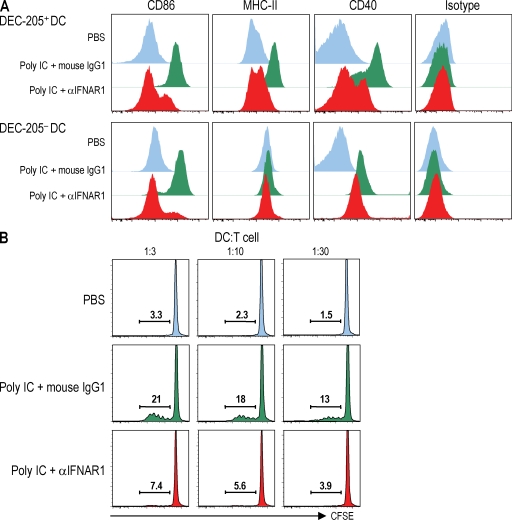
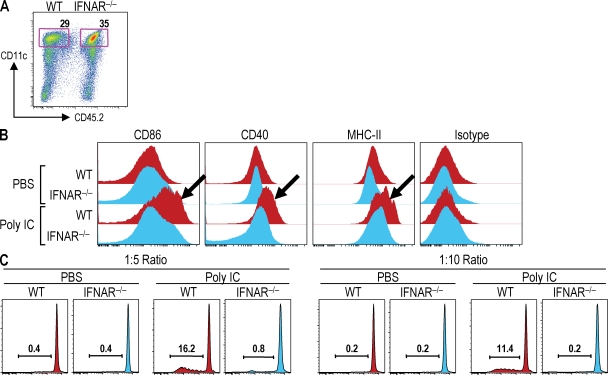
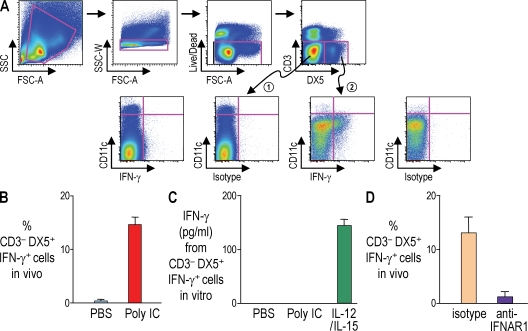
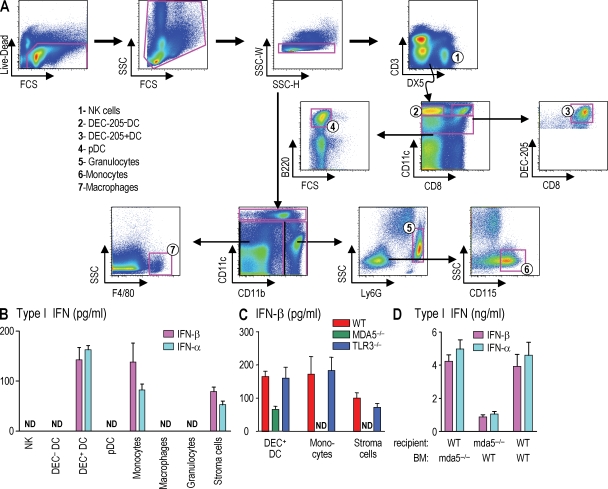
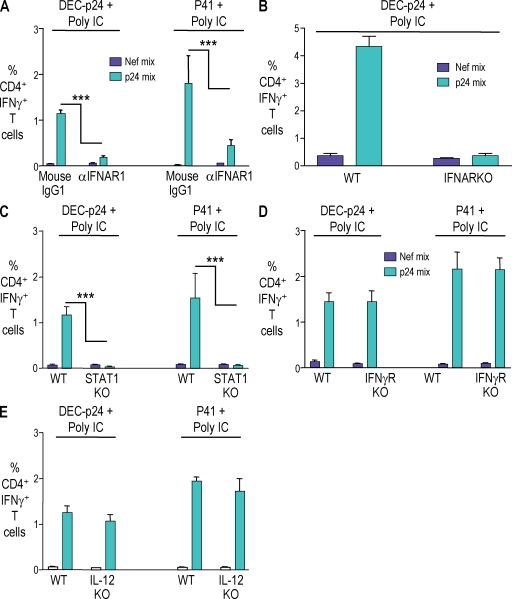
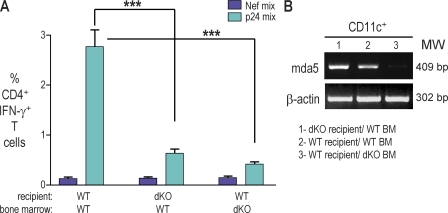
Relative to several other toll-like receptor (TLR) agonists, we found polyinosinic:polycytidylic acid (poly IC) to be the most effective adjuvant for Th1 CD4(+) T cell responses to a dendritic cell (DC)-targeted HIV gag protein vaccine in mice. To identify mechanisms for adjuvant action in the intact animal and the polyclonal T cell repertoire, we found poly IC to be the most effective inducer of type I interferon (IFN), which was produced by DEC-205(+) DCs, monocytes, and stromal cells. Antibody blocking or deletion of type I IFN receptor showed that IFN was essential for DC maturation and development of CD4(+) immunity. The IFN-AR receptor was directly required for DCs to respond to poly IC. STAT 1 was also essential, in keeping with the type I IFN requirement, but not type II IFN or IL-12 p40. Induction of type I IFN was mda5 dependent, but DCs additionally used TLR3. In bone marrow chimeras, radioresistant and, likely, nonhematopoietic cells were the main source of IFN, but mda5 was required in both marrow-derived and radioresistant host cells for adaptive responses. Therefore, the adjuvant action of poly IC requires a widespread innate type I IFN response that directly links antigen presentation by DCs to adaptive immunity.
Figures
Similar articles
-
The microbial mimic poly IC induces durable and protective CD4+ T cell immunity together with a dendritic cell targeted vaccine.Proc Natl Acad Sci U S A. 2008 Feb 19;105(7):2574-9. doi: 10.1073/pnas.0711976105. Epub 2008 Feb 6. Proc Natl Acad Sci U S A. 2008. PMID: 18256187 Free PMC article.
-
The combination of early and rapid type I IFN, IL-1α, and IL-1β production are essential mediators of RNA-like adjuvant driven CD4+ Th1 responses.PLoS One. 2011;6(12):e29412. doi: 10.1371/journal.pone.0029412. Epub 2011 Dec 19. PLoS One. 2011. PMID: 22206014 Free PMC article.
-
Type I interferon is selectively required by dendritic cells for immune rejection of tumors.J Exp Med. 2011 Sep 26;208(10):1989-2003. doi: 10.1084/jem.20101158. Epub 2011 Sep 19. J Exp Med. 2011. PMID: 21930769 Free PMC article.
-
Particulate formulations for the delivery of poly(I:C) as vaccine adjuvant.Adv Drug Deliv Rev. 2013 Oct;65(10):1386-99. doi: 10.1016/j.addr.2013.05.013. Epub 2013 Jun 7. Adv Drug Deliv Rev. 2013. PMID: 23751781 Review.
-
Two Modes of Th1 Polarization Induced by Dendritic-Cell-Priming Adjuvant in Vaccination.Cells. 2023 May 29;12(11):1504. doi: 10.3390/cells12111504. Cells. 2023. PMID: 37296625 Free PMC article. Review.
Cited by
-
The Regulation of Immune Responses by DC Derived Type I IFN.Front Immunol. 2013 Apr 22;4:94. doi: 10.3389/fimmu.2013.00094. eCollection 2013. Front Immunol. 2013. PMID: 23626590 Free PMC article.
-
Targeting Leishmania major Antigens to Dendritic Cells In Vivo Induces Protective Immunity.PLoS One. 2013 Jun 26;8(6):e67453. doi: 10.1371/journal.pone.0067453. Print 2013. PLoS One. 2013. PMID: 23840706 Free PMC article.
-
A microarray patch SARS-CoV-2 vaccine induces sustained antibody responses and polyfunctional cellular immunity.iScience. 2022 Oct 21;25(10):105045. doi: 10.1016/j.isci.2022.105045. Epub 2022 Aug 30. iScience. 2022. PMID: 36062075 Free PMC article.
-
Combined adjuvants of poly(I:C) plus LAG-3-Ig improve antitumor effects of tumor-specific T cells, preventing their exhaustion.Cancer Sci. 2016 Apr;107(4):398-406. doi: 10.1111/cas.12861. Cancer Sci. 2016. PMID: 27079438 Free PMC article.
-
Potent Cell-Intrinsic Immune Responses in Dendritic Cells Facilitate HIV-1-Specific T Cell Immunity in HIV-1 Elite Controllers.PLoS Pathog. 2015 Jun 11;11(6):e1004930. doi: 10.1371/journal.ppat.1004930. eCollection 2015 Jun. PLoS Pathog. 2015. PMID: 26067651 Free PMC article.
References
-
- Afkarian M., Sedy J.R., Yang J., Jacobson N.G., Cereb N., Yang S.Y., Murphy T.L., Murphy K.M. 2002. T-bet is a STAT1-induced regulator of IL-12R expression in naive CD4+ T cells.Nat. Immunol. 3:549–557 - PubMed
-
- Akira S., Uematsu S., Takeuchi O. 2006. Pathogen recognition and innate immunity.Cell. 124:783–801 - PubMed
-
- Alexopoulou L., Holt A.C., Medzhitov R., Flavell R.A. 2001. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3.Nature. 413:732–738 - PubMed
-
- Berenson L.S., Farrar J.D., Murphy T.L., Murphy K.M. 2004. Frontline: absence of functional STAT4 activation despite detectable tyrosine phosphorylation induced by murine IFN-α.Eur. J. Immunol. 34:2365–2374 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
