

Inflammasome-mediated disease animal models reveal roles for innate but not adaptive immunity
- PMID: 19501000
- PMCID: PMC2759865
- DOI: 10.1016/j.immuni.2009.05.005
Inflammasome-mediated disease animal models reveal roles for innate but not adaptive immunity
Abstract
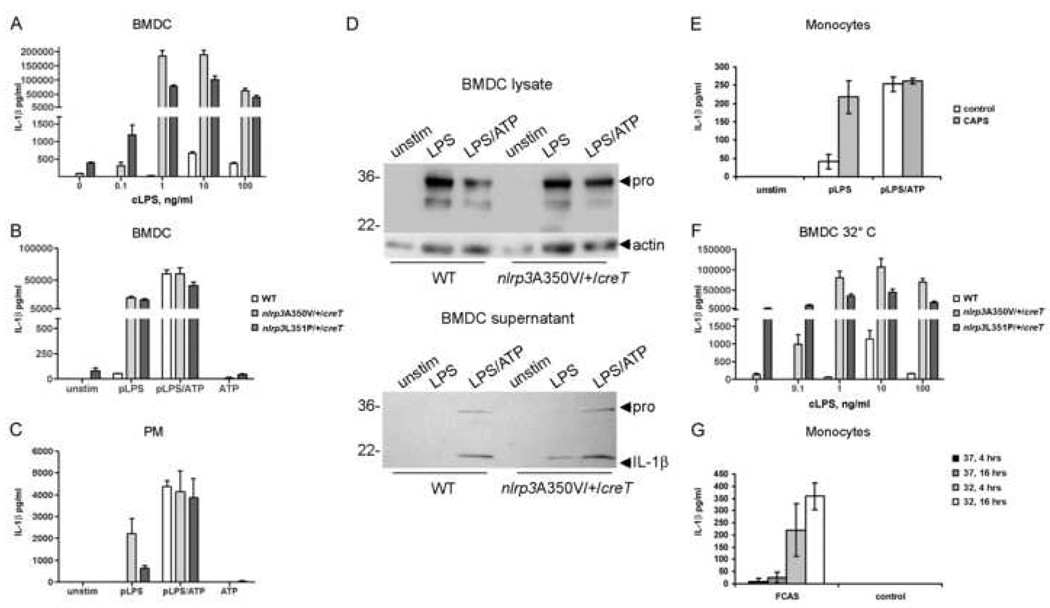
NLRP3 nucleates the inflammasome, a protein complex responsible for cleavage of prointerleukin-1beta (IL-1beta) to its active form. Mutations in the NLRP3 gene cause the autoinflammatory disease spectrum cryopyrin-associated periodic syndromes (CAPS). The central role of IL-1beta in CAPS is supported by the response to IL-1-targeted therapy. We developed two Nlrp3 mutant knockin mouse strains to model CAPS to examine the role of other inflammatory mediators and adaptive immune responses in an innate immune-driven disease. These mice had systemic inflammation and poor growth, similar to some human CAPS patients, and demonstrated early mortality, primarily mediated by myeloid cells. Mating these mutant mice to various gene mutant backgrounds showed that the mouse disease phenotype required an intact inflammasome, was only partially dependent on IL-1beta, and was independent of T cells. These data suggest that CAPS are true inflammasome-mediated diseases and provide insight for more common inflammatory disorders.
Figures





Comment in
-
Knocking in the NLRP3 inflammasome.Immunity. 2009 Jun 19;30(6):761-3. doi: 10.1016/j.immuni.2009.06.001. Immunity. 2009. PMID: 19538926
Similar articles
-
Divergence of IL-1, IL-18, and cell death in NLRP3 inflammasomopathies.J Clin Invest. 2013 Nov;123(11):4695-705. doi: 10.1172/JCI71543. J Clin Invest. 2013. PMID: 24084736 Free PMC article.
-
A novel knock-in mouse model of cryopyrin-associated periodic syndromes with development of amyloidosis: Therapeutic efficacy of proton pump inhibitors.J Allergy Clin Immunol. 2020 Jan;145(1):368-378.e13. doi: 10.1016/j.jaci.2019.05.034. Epub 2019 Jun 10. J Allergy Clin Immunol. 2020. PMID: 31194989
-
Knocking in the NLRP3 inflammasome.Immunity. 2009 Jun 19;30(6):761-3. doi: 10.1016/j.immuni.2009.06.001. Immunity. 2009. PMID: 19538926
-
The inflammasome, autoinflammatory diseases, and gout.Joint Bone Spine. 2007 Dec;74(6):571-6. doi: 10.1016/j.jbspin.2007.04.004. Epub 2007 Aug 21. Joint Bone Spine. 2007. PMID: 17714972 Review.
-
The discovery of NLRP3 and its function in cryopyrin-associated periodic syndromes and innate immunity.Immunol Rev. 2024 Mar;322(1):259-282. doi: 10.1111/imr.13292. Epub 2023 Dec 25. Immunol Rev. 2024. PMID: 38146057 Review.
Cited by
-
Cutting edge: nitrogen bisphosphonate-induced inflammation is dependent upon mast cells and IL-1.J Immunol. 2012 Apr 1;188(7):2977-80. doi: 10.4049/jimmunol.1100830. Epub 2012 Mar 2. J Immunol. 2012. PMID: 22387558 Free PMC article.
-
Equally potent: Nlrp3 mutation in macrophages or neutrophils is sufficient to drive autoinflammation.EMBO Rep. 2022 Nov 7;23(11):e56091. doi: 10.15252/embr.202256091. Epub 2022 Oct 4. EMBO Rep. 2022. PMID: 36194522 Free PMC article.
-
G-CSF drives autoinflammation in APLAID.Nat Immunol. 2023 May;24(5):814-826. doi: 10.1038/s41590-023-01473-6. Epub 2023 Mar 30. Nat Immunol. 2023. PMID: 36997670 Free PMC article.
-
Potential of IL-1, IL-18 and Inflammasome Inhibition for the Treatment of Inflammatory Skin Diseases.Front Pharmacol. 2017 May 22;8:278. doi: 10.3389/fphar.2017.00278. eCollection 2017. Front Pharmacol. 2017. PMID: 28588486 Free PMC article. Review.
-
Hepatocyte pyroptosis and release of inflammasome particles induce stellate cell activation and liver fibrosis.J Hepatol. 2021 Jan;74(1):156-167. doi: 10.1016/j.jhep.2020.07.041. Epub 2020 Aug 4. J Hepatol. 2021. PMID: 32763266 Free PMC article.
References
-
- Agostini L, Martinon F, Burns K, McDermott MF, Hawkins PN, Tschopp J. NALP3 forms an IL-1beta-processing inflammasome with increased activity in Muckle-Wells autoinflammatory disorder. Immunity. 2004;20:319–325. - PubMed
-
- Barnden MJ, Allison J, Heath WR, Carbone FR. Defective TCR expression in transgenic mice constructed using cDNA-based alpha- and beta-chain genes under the control of heterologous regulatory elements. Immunol Cell Biol. 1998;76:34–40. - PubMed
-
- Chae JJ, Komarow HD, Cheng J, Wood G, Raben N, Liu PP, Kastner DL. Targeted disruption of pyrin, the FMF protein, causes heightened sensitivity to endotoxin and a defect in macrophage apoptosis. Mol Cell. 2003;11:591–604. - PubMed
-
- Clausen BE, Burkhardt C, Reith W, Renkawitz R, Forster I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res. 1999;8:265–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
