

Tumor vasculature is regulated by PHD2-mediated angiogenesis and bone marrow-derived cell recruitment
- PMID: 19477431
- PMCID: PMC2846696
- DOI: 10.1016/j.ccr.2009.04.010
Tumor vasculature is regulated by PHD2-mediated angiogenesis and bone marrow-derived cell recruitment
Abstract
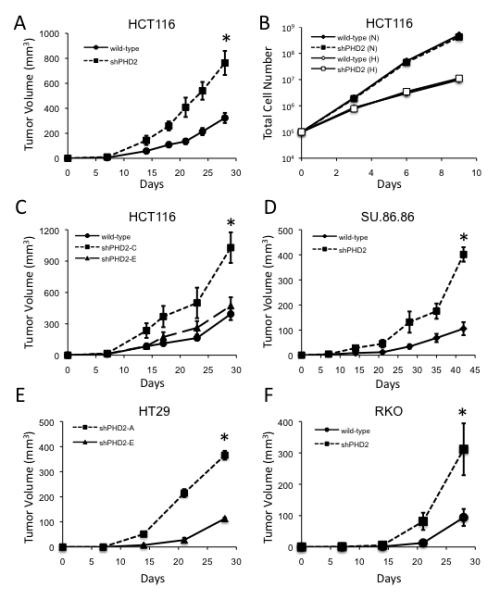
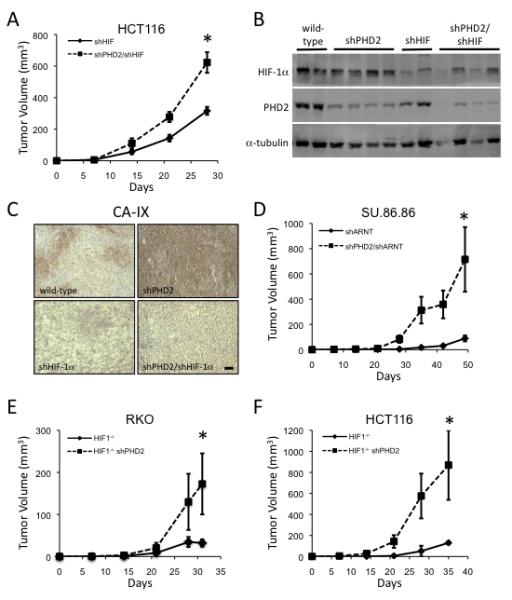
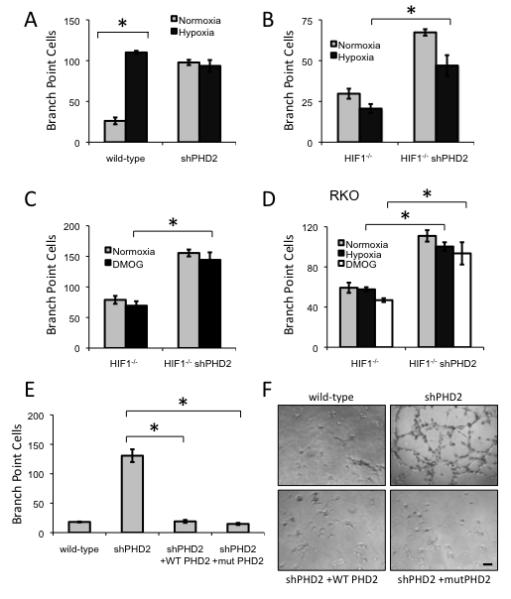
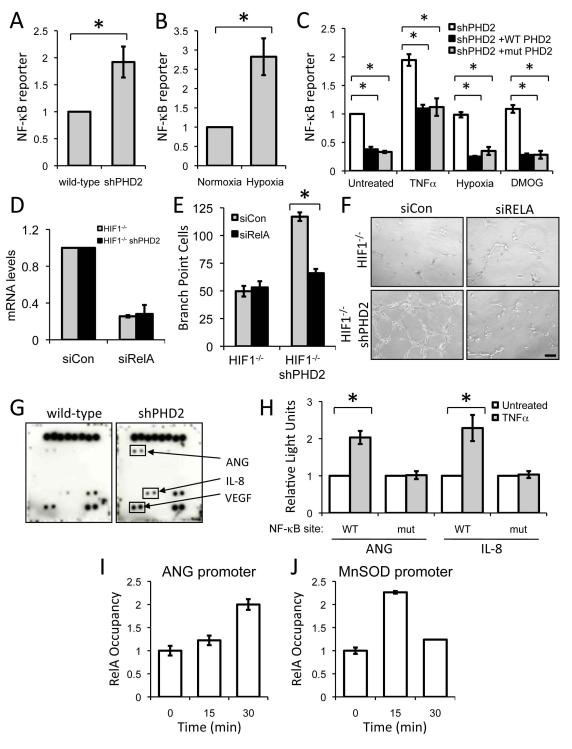
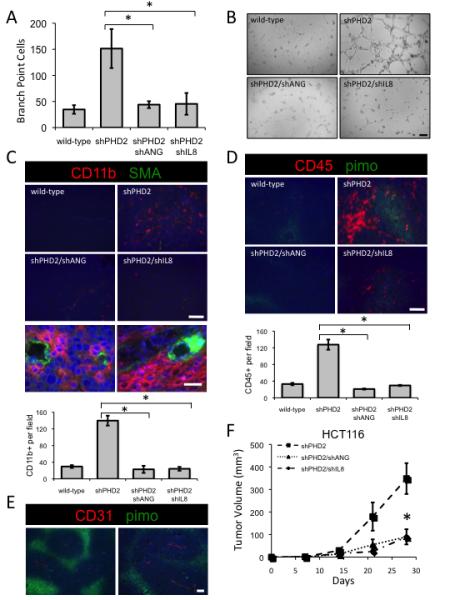
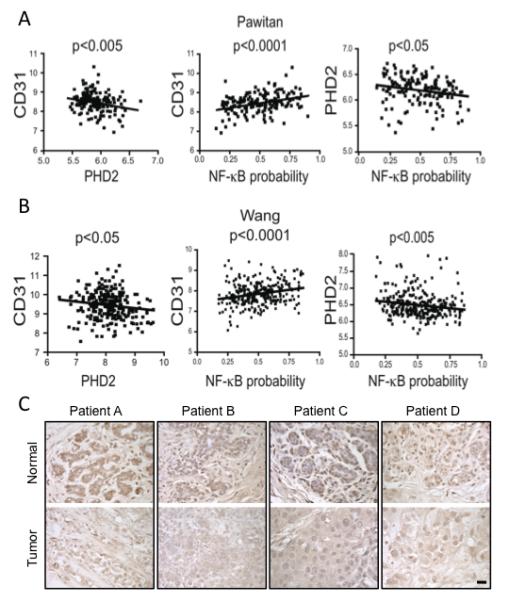
Sustained angiogenesis, through either local sprouting (angiogenesis) or the recruitment of bone marrow-derived cells (BMDCs) (vasculogenesis), is essential to the development of a tumor. How BMDCs are recruited to the tumor and their contribution to the tumor vasculature is poorly understood. Here, we demonstrate that both IL-8 and angiogenin contribute to the complementary pathways of angiogenesis and BMDC mobilization to increase tumor growth. These two factors are regulated by PHD2 in a HIF-independent but NF-kappaB-dependent manner. PHD2 levels are decreased in human cancers, compared with corresponding normal tissue, and correlate with an increase in mature blood vessels. Thus, PHD2 plays a critical role in regulating tumor angiogenesis.
Figures


Similar articles
-
PHD2 in tumour angiogenesis.Br J Cancer. 2010 Jun 29;103(1):1-5. doi: 10.1038/sj.bjc.6605682. Epub 2010 May 11. Br J Cancer. 2010. PMID: 20461086 Free PMC article. Review.
-
Prolyl hydroxylase-2 (PHD2) exerts tumor-suppressive activity in pancreatic cancer.Cancer. 2012 Feb 15;118(4):960-72. doi: 10.1002/cncr.26344. Epub 2011 Jul 26. Cancer. 2012. PMID: 21792862
-
Vascular endothelial growth factor (VEGF) modulation by targeting hypoxia-inducible factor-1alpha--> hypoxia response element--> VEGF cascade differentially regulates vascular response and growth rate in tumors.Cancer Res. 2000 Nov 15;60(22):6248-52. Cancer Res. 2000. PMID: 11103778
-
Lactate influx through the endothelial cell monocarboxylate transporter MCT1 supports an NF-κB/IL-8 pathway that drives tumor angiogenesis.Cancer Res. 2011 Apr 1;71(7):2550-60. doi: 10.1158/0008-5472.CAN-10-2828. Epub 2011 Feb 7. Cancer Res. 2011. PMID: 21300765
-
[Hypoxia-responsive factor PHD2 and angiogenic diseases].Yao Xue Xue Bao. 2014 Feb;49(2):151-7. Yao Xue Xue Bao. 2014. PMID: 24761603 Review. Chinese.
Cited by
-
PHD2: from hypoxia regulation to disease progression.Hypoxia (Auckl). 2016 Apr 11;4:53-67. doi: 10.2147/HP.S53576. eCollection 2016. Hypoxia (Auckl). 2016. PMID: 27800508 Free PMC article. Review.
-
Endothelin-1 inhibits prolyl hydroxylase domain 2 to activate hypoxia-inducible factor-1alpha in melanoma cells.PLoS One. 2010 Jun 21;5(6):e11241. doi: 10.1371/journal.pone.0011241. PLoS One. 2010. Retraction in: PLoS One. 2024 Dec 5;19(12):e0315282. doi: 10.1371/journal.pone.0315282 PMID: 20574527 Free PMC article. Retracted.
-
Human bone marrow mesenchymal stem cells display anti-cancer activity in SCID mice bearing disseminated non-Hodgkin's lymphoma xenografts.PLoS One. 2010 Jun 16;5(6):e11140. doi: 10.1371/journal.pone.0011140. PLoS One. 2010. PMID: 20585401 Free PMC article.
-
The oxygen sensor prolyl hydroxylase domain 2 regulates the in vivo suppressive capacity of regulatory T cells.Elife. 2022 Feb 22;11:e70555. doi: 10.7554/eLife.70555. Elife. 2022. PMID: 35192456 Free PMC article.
-
WSB-1 regulates the metastatic potential of hormone receptor negative breast cancer.Br J Cancer. 2018 May;118(9):1229-1237. doi: 10.1038/s41416-018-0056-3. Epub 2018 Mar 15. Br J Cancer. 2018. PMID: 29540773 Free PMC article.
References
-
- Appelhoff RJ, Tian YM, Raval RR, Turley H, Harris AL, Pugh CW, Ratcliffe PJ, Gleadle JM. Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J Biol Chem. 2004;279:38458–38465. - PubMed
-
- Bild AH, Yao G, Chang JT, Wang Q, Potti A, Chasse D, Joshi MB, Harpole D, Lancaster JM, Berchuck A, et al. Oncogenic pathway signatures in human cancers as a guide to targeted therapies. Nature. 2006;439:353–357. - PubMed
-
- Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat Med. 2000;6:389–395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32 CA123823-02/CA/NCI NIH HHS/United States
- NCI-CA-123823/CA/NCI NIH HHS/United States
- P01 CA067166/CA/NCI NIH HHS/United States
- CA-125618/CA/NCI NIH HHS/United States
- NCI-CA-67166/CA/NCI NIH HHS/United States
- F32 CA123823/CA/NCI NIH HHS/United States
- F32 CA123823-01/CA/NCI NIH HHS/United States
- T32 CA009302/CA/NCI NIH HHS/United States
- R01 CA116685/CA/NCI NIH HHS/United States
- R01 CA125618/CA/NCI NIH HHS/United States
- F32 CA123823-03/CA/NCI NIH HHS/United States
- NCI-CA-116685/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
