

The effect of ligand efficacy on the formation and stability of a GPCR-G protein complex
- PMID: 19470481
- PMCID: PMC2685739
- DOI: 10.1073/pnas.0811437106
The effect of ligand efficacy on the formation and stability of a GPCR-G protein complex
Abstract
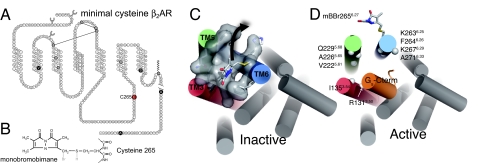
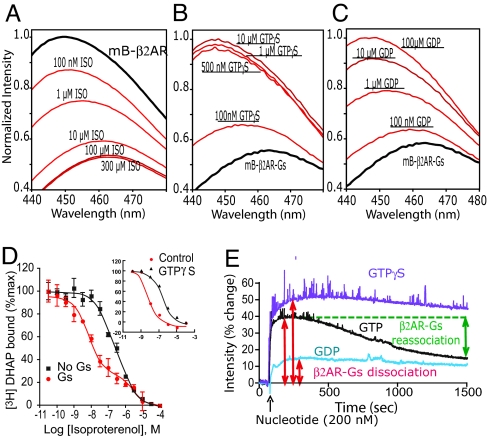
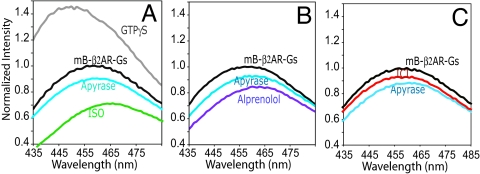
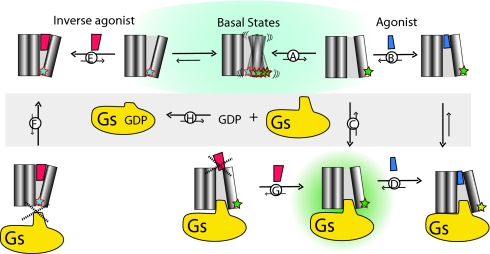
G protein-coupled receptors (GPCRs) mediate the majority of physiologic responses to hormones and neurotransmitters. However, many GPCRs exhibit varying degrees of agonist-independent G protein activation. This phenomenon is referred to as basal or constitutive activity. For many of these GPCRs, drugs classified as inverse agonists can suppress basal activity. There is a growing body of evidence that basal activity is physiologically relevant, and the ability of a drug to inhibit basal activity may influence its therapeutic properties. However, the molecular mechanism for basal activation and inhibition of basal activity by inverse agonists is poorly understood and difficult to study, because the basally active state is short-lived and represents a minor fraction of receptor conformations. Here, we investigate basal activation of the G protein Gs by the beta(2) adrenergic receptor (beta(2)AR) by using purified receptor reconstituted into recombinant HDL particles with a stoichiometric excess of Gs. The beta(2)AR is site-specifically labeled with a small, environmentally sensitive fluorophore enabling direct monitoring of agonist- and Gs-induced conformational changes. In the absence of an agonist, the beta(2)AR and Gs can be trapped in a complex by enzymatic depletion of guanine nucleotides. Formation of the complex is enhanced by the agonist isoproterenol, and it rapidly dissociates on exposure to concentrations of GTP and GDP found in the cytoplasm. The inverse agonist ICI prevents formation of the beta(2)AR-Gs complex, but has little effect on preformed complexes. These results provide insights into G protein-induced conformational changes in the beta(2)AR and the structural basis for ligand efficacy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Allosteric coupling from G protein to the agonist-binding pocket in GPCRs.Nature. 2016 Jul 7;535(7610):182-6. doi: 10.1038/nature18324. Epub 2016 Jun 29. Nature. 2016. PMID: 27362234 Free PMC article.
-
Functional differences between full and partial agonists: evidence for ligand-specific receptor conformations.J Pharmacol Exp Ther. 2001 Jun;297(3):1218-26. J Pharmacol Exp Ther. 2001. PMID: 11356949
-
Detection of G protein-selective G protein-coupled receptor (GPCR) conformations in live cells.J Biol Chem. 2013 Jun 14;288(24):17167-78. doi: 10.1074/jbc.M113.464065. Epub 2013 Apr 29. J Biol Chem. 2013. PMID: 23629648 Free PMC article.
-
G protein-coupled receptors--recent advances.Acta Biochim Pol. 2012;59(4):515-29. Epub 2012 Dec 18. Acta Biochim Pol. 2012. PMID: 23251911 Free PMC article. Review.
-
Structural features of β2 adrenergic receptor: crystal structures and beyond.Mol Cells. 2015;38(2):105-11. doi: 10.14348/molcells.2015.2301. Epub 2014 Dec 24. Mol Cells. 2015. PMID: 25537861 Free PMC article. Review.
Cited by
-
Positive allosteric modulation of a GPCR ternary complex.Sci Adv. 2024 Sep 13;10(37):eadp7040. doi: 10.1126/sciadv.adp7040. Epub 2024 Sep 11. Sci Adv. 2024. PMID: 39259792 Free PMC article.
-
Agonist-induced formation of unproductive receptor-G12 complexes.Proc Natl Acad Sci U S A. 2020 Sep 1;117(35):21723-21730. doi: 10.1073/pnas.2003787117. Epub 2020 Aug 17. Proc Natl Acad Sci U S A. 2020. PMID: 32817560 Free PMC article.
-
Therapeutic antibodies directed at G protein-coupled receptors.MAbs. 2010 Nov-Dec;2(6):594-606. doi: 10.4161/mabs.2.6.13420. Epub 2010 Nov 1. MAbs. 2010. PMID: 20864805 Free PMC article. Review.
-
Computational design of G Protein-Coupled Receptor allosteric signal transductions.Nat Chem Biol. 2020 Jan;16(1):77-86. doi: 10.1038/s41589-019-0407-2. Epub 2019 Dec 2. Nat Chem Biol. 2020. PMID: 31792443
-
Stability of the neurotensin receptor NTS1 free in detergent solution and immobilized to affinity resin.PLoS One. 2010 Sep 7;5(9):e12579. doi: 10.1371/journal.pone.0012579. PLoS One. 2010. PMID: 20830205 Free PMC article.
References
-
- Lefkowitz RJ, Shenoy SK. Transduction of receptor signals by beta-arrestins. Science. 2005;308:512–517. - PubMed
-
- Luttrell LM, Lefkowitz RJ. The role of beta-arrestins in the termination and transduction of G-protein-coupled receptor signals. J Cell Sci. 2002;115:455–465. - PubMed
-
- Sunahara RK, Tesmer JJ, Gilman AG, Sprang SR. Crystal structure of the adenylyl cyclase activator Gsα. Science. 1997;278:1943–1947. - PubMed
-
- Sondek J, Lambright DG, Noel JP, Hamm HE, Sigler PB. GTPase mechanism of G-proteins from the 1.7-Å crystal structure of transducin α-GDP AIF−4. Nature. 1994;372:276–279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
