

Ubiquitin-mediated degradation of the formin mDia2 upon completion of cell division
- PMID: 19457867
- PMCID: PMC2740432
- DOI: 10.1074/jbc.M109.000885
Ubiquitin-mediated degradation of the formin mDia2 upon completion of cell division
Abstract
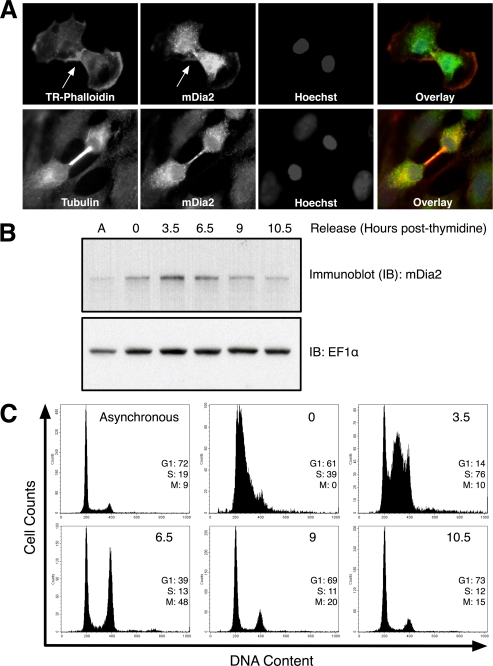
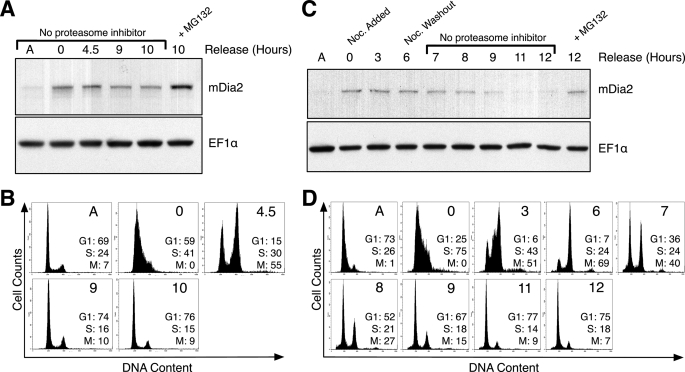
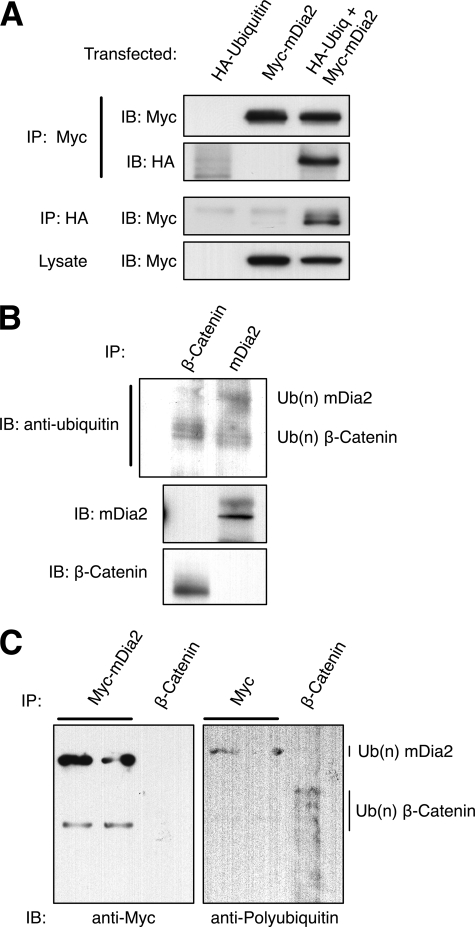
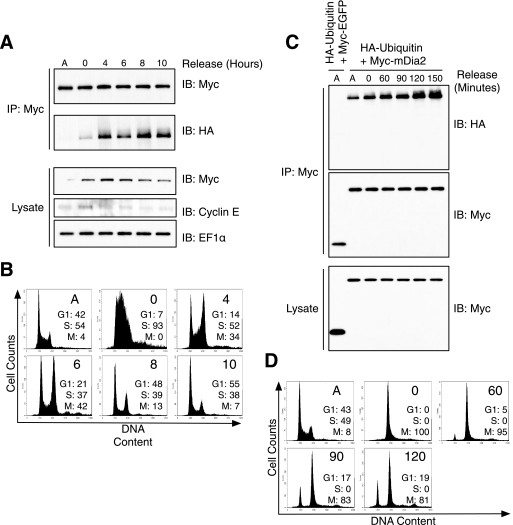
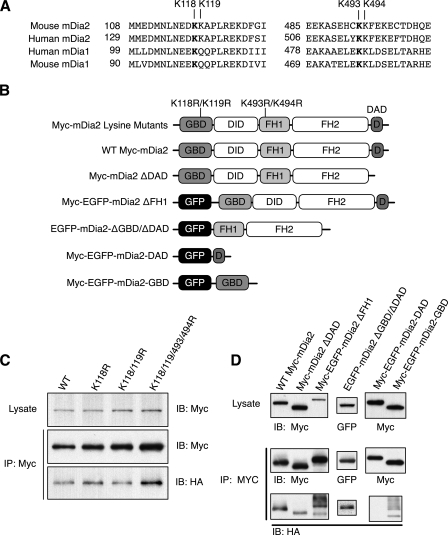
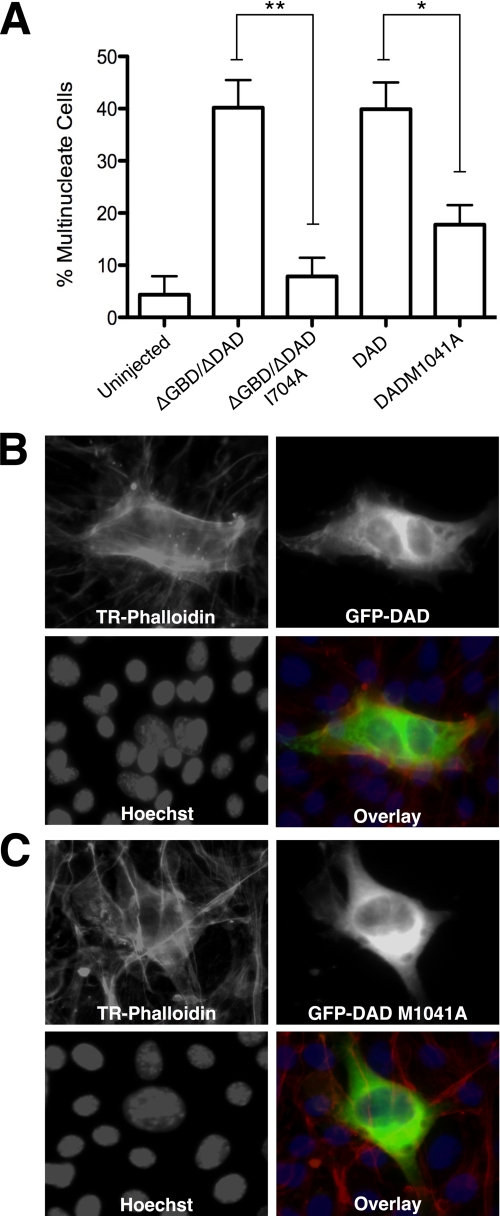
Formins assemble non-branched actin filaments and modulate microtubule dynamics during cell migration and cell division. At the end of mitosis formins contribute to the generation of actin filaments that form the contractile ring. Rho small GTP-binding proteins activate mammalian diaphanous-related (mDia) formins by directly binding and disrupting an intramolecular autoinhibitory mechanism. Although the Rho-regulated activation mechanism is well characterized, little is known about how formins are switched off. Here we reveal a novel mechanism of formin regulation during cytokinesis based on the following observations; 1) mDia2 is degraded at the end of mitosis, 2) mDia2 is targeted for disposal by post-translational ubiquitin modification, 3) forced expression of activated mDia2 yields binucleate cells due to failed cytokinesis, and 4) the cytokinesis block is dependent upon mDia2-mediated actin assembly as versions of mDia2 incapable of nucleating actin but that still stabilize microtubules have no effect on cytokinesis. We propose that the tight control of mDia2 expression and ubiquitin-mediated degradation is essential for the completion of cell division. Because of the many roles for formins in cell morphology, we discuss the relevance of mDia protein turnover in other processes where ubiquitin-mediated proteolysis is an essential component.
Figures
Similar articles
-
Dia-interacting protein modulates formin-mediated actin assembly at the cell cortex.Curr Biol. 2007 Apr 3;17(7):579-91. doi: 10.1016/j.cub.2007.03.024. Curr Biol. 2007. PMID: 17398099
-
Rho activation of mDia formins is modulated by an interaction with inverted formin 2 (INF2).Proc Natl Acad Sci U S A. 2011 Feb 15;108(7):2933-8. doi: 10.1073/pnas.1017010108. Epub 2011 Jan 28. Proc Natl Acad Sci U S A. 2011. PMID: 21278336 Free PMC article.
-
Novel roles of formin mDia2 in lamellipodia and filopodia formation in motile cells.PLoS Biol. 2007 Nov;5(11):e317. doi: 10.1371/journal.pbio.0050317. PLoS Biol. 2007. PMID: 18044991 Free PMC article.
-
Mechanism and function of formins in the control of actin assembly.Annu Rev Biochem. 2007;76:593-627. doi: 10.1146/annurev.biochem.75.103004.142647. Annu Rev Biochem. 2007. PMID: 17373907 Review.
-
The role of formins in human disease.Biochim Biophys Acta. 2010 Feb;1803(2):226-33. doi: 10.1016/j.bbamcr.2009.11.006. Epub 2009 Nov 24. Biochim Biophys Acta. 2010. PMID: 19941910 Review.
Cited by
-
The Insulin Receptor Adaptor IRS2 is an APC/C Substrate That Promotes Cell Cycle Protein Expression and a Robust Spindle Assembly Checkpoint.Mol Cell Proteomics. 2020 Sep;19(9):1450-1467. doi: 10.1074/mcp.RA120.002069. Epub 2020 Jun 18. Mol Cell Proteomics. 2020. PMID: 32554797 Free PMC article.
-
Direct interaction between two actin nucleators is required in Drosophila oogenesis.Development. 2013 Nov;140(21):4417-25. doi: 10.1242/dev.097337. Epub 2013 Oct 2. Development. 2013. PMID: 24089467 Free PMC article.
-
Executable Network Models of Integrated Multiomics Data.J Proteome Res. 2023 May 5;22(5):1546-1556. doi: 10.1021/acs.jproteome.2c00730. Epub 2023 Mar 31. J Proteome Res. 2023. PMID: 37000949 Free PMC article.
-
Stress-activated MAPK signaling controls fission yeast actomyosin ring integrity by modulating formin For3 levels.Elife. 2020 Sep 11;9:e57951. doi: 10.7554/eLife.57951. Elife. 2020. PMID: 32915139 Free PMC article.
-
Astrocytes control quiescent NSC reactivation via GPCR signaling-mediated F-actin remodeling.Sci Adv. 2024 Jul 26;10(30):eadl4694. doi: 10.1126/sciadv.adl4694. Epub 2024 Jul 24. Sci Adv. 2024. PMID: 39047090 Free PMC article.
References
-
- Eisenmann K. M., West R. A., Hildebrand D., Kitchen S. M., Peng J., Sigler R., Zhang J., Siminovitch K. A., Alberts A. S. (2007) J. Biol. Chem. 282, 25152–25158 - PubMed
-
- Gupton S. L., Eisenmann K., Alberts A. S., Waterman-Storer C. M. (2007) J. Cell Sci. 120, 3475–3487 - PubMed
-
- Wallar B. J., Deward A. D., Resau J. H., Alberts A. S. (2007) Exp. Cell Res. 313, 560–571 - PubMed
-
- Fernandez-Borja M., Janssen L., Verwoerd D., Hordijk P., Neefjes J. (2005) J. Cell Sci. 118, 2661–2670 - PubMed
-
- Peng J., Kitchen S. M., West R. A., Sigler R., Eisenmann K. M., Alberts A. S. (2007) Cancer Res. 67, 7565–7571 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
