

An inhibitory interaction between viral and cellular proteins underlies the resistance of tomato to nonadapted tobamoviruses
- PMID: 19423673
- PMCID: PMC2678887
- DOI: 10.1073/pnas.0809105106
An inhibitory interaction between viral and cellular proteins underlies the resistance of tomato to nonadapted tobamoviruses
Abstract
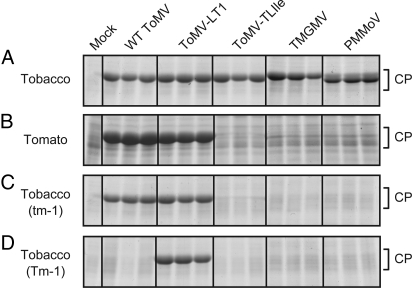
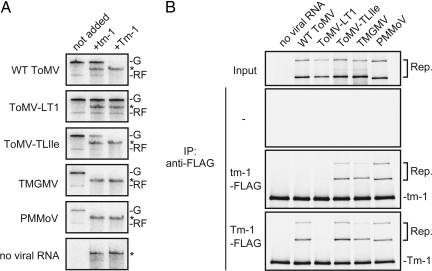
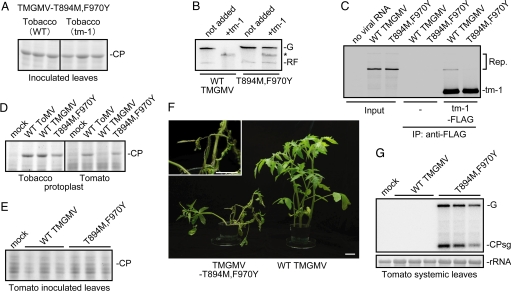
Any individual virus can infect only a limited range of hosts, and most plant species are "nonhosts" to a given virus; i.e., all members of the species are insusceptible to the virus. In nonhost plants, the factors that control virus resistance are not genetically tractable, and how the host range of a virus is determined remains poorly understood. Tomato (Solanum lycopersicum) is a nonhost species for Tobacco mild green mosaic virus (TMGMV) and Pepper mild mottle virus (PMMoV), members of the genus Tobamovirus. Previously, we identified Tm-1, a resistance gene of tomato to another tobamovirus, Tomato mosaic virus (ToMV), and found that Tm-1 binds to ToMV replication proteins to inhibit RNA replication. Tm-1 is derived from a wild tomato species, S. habrochaites, and ToMV-susceptible tomato cultivars have the allelic gene tm-1. The tm-1 protein can neither bind to ToMV replication proteins nor inhibit ToMV multiplication. Here, we show that transgenic tobacco plants expressing tm-1 exhibit resistance to TMGMV and PMMoV. The tm-1 protein bound to the replication proteins of TMGMV and PMMoV and inhibited their RNA replication in vitro. In one of the tm-1-expressing tobacco plants, a tm-1-insensitive TMGMV mutant emerged. In tomato protoplasts, this mutant TMGMV multiplied as efficiently as ToMV. However, in tomato plants, the mutant TMGMV multiplied with lower efficiency compared to ToMV and caused systemic necrosis. These results suggest that an inhibitory interaction between the replication proteins and tm-1 underlies a multilayered resistance mechanism to TMGMV in tomato.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Coevolution and hierarchical interactions of Tomato mosaic virus and the resistance gene Tm-1.PLoS Pathog. 2012;8(10):e1002975. doi: 10.1371/journal.ppat.1002975. Epub 2012 Oct 18. PLoS Pathog. 2012. PMID: 23093939 Free PMC article.
-
Gaining replicability in a nonhost compromises the silencing suppression activity of Tobacco mild green mosaic virus in a host.J Virol. 2011 Feb;85(4):1893-5. doi: 10.1128/JVI.01741-10. Epub 2010 Nov 24. J Virol. 2011. PMID: 21106731 Free PMC article.
-
Detection of tobamoviruses by RT-PCR using a novel pair of degenerate primers.J Virol Methods. 2018 Sep;259:122-128. doi: 10.1016/j.jviromet.2018.06.012. Epub 2018 Jun 23. J Virol Methods. 2018. PMID: 29944907
-
Mechanisms of tomato mosaic virus RNA replication and its inhibition by the host resistance factor Tm-1.Curr Opin Virol. 2014 Dec;9:8-13. doi: 10.1016/j.coviro.2014.08.005. Epub 2014 Sep 17. Curr Opin Virol. 2014. PMID: 25212767 Review.
-
Engineered Resistance to Tobamoviruses.Viruses. 2024 Jun 22;16(7):1007. doi: 10.3390/v16071007. Viruses. 2024. PMID: 39066170 Free PMC article. Review.
Cited by
-
Movement Protein Mediates Systemic Necrosis in Tomato Plants with Infection of Tomato Mosaic Virus.Viruses. 2023 Jan 4;15(1):157. doi: 10.3390/v15010157. Viruses. 2023. PMID: 36680197 Free PMC article.
-
ARGONAUTE2 mediates RNA-silencing antiviral defenses against Potato virus X in Arabidopsis.Plant Physiol. 2011 Jul;156(3):1556-64. doi: 10.1104/pp.111.178012. Epub 2011 May 16. Plant Physiol. 2011. PMID: 21576511 Free PMC article.
-
A spotlight on non-host resistance to plant viruses.PeerJ. 2022 Mar 31;10:e12996. doi: 10.7717/peerj.12996. eCollection 2022. PeerJ. 2022. PMID: 35382007 Free PMC article.
-
Functional characterization of the mutations in Pepper mild mottle virus overcoming tomato tm-1-mediated resistance.Mol Plant Pathol. 2014 Jun;15(5):479-87. doi: 10.1111/mpp.12107. Epub 2014 Jan 19. Mol Plant Pathol. 2014. PMID: 24299004 Free PMC article.
-
Coevolution and hierarchical interactions of Tomato mosaic virus and the resistance gene Tm-1.PLoS Pathog. 2012;8(10):e1002975. doi: 10.1371/journal.ppat.1002975. Epub 2012 Oct 18. PLoS Pathog. 2012. PMID: 23093939 Free PMC article.
References
-
- Kang BC, Yeam I, Jahn MM. Genetics of plant virus resistance. Annu Rev Phytopathol. 2005;43:581–621. - PubMed
-
- Maule AJ, Caranta C, Boulton MI. Sources of natural resistance to plant viruses: status and prospects. Mol Plant Pathol. 2007;8:223–231. - PubMed
-
- Soosaar JLM, Burch-Smith TM, Dinesh-Kumar SP. Mechanisms of plant resistance to viruses. Nat Rev Microbiol. 2005;3:789–798. - PubMed
-
- Diaz-Pendon JA, et al. Advances in understanding recessive resistance to plant viruses. Mol Plant Pathol. 2004;5:223–233. - PubMed
-
- Robaglia C, Caranta C. Translation initiation factors: A weak link in plant RNA virus infection. Trends Plant Sci. 2006;11:40–45. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
