

Critical regulation of early Th17 cell differentiation by interleukin-1 signaling
- PMID: 19362022
- PMCID: PMC2705871
- DOI: 10.1016/j.immuni.2009.02.007
Critical regulation of early Th17 cell differentiation by interleukin-1 signaling
Abstract
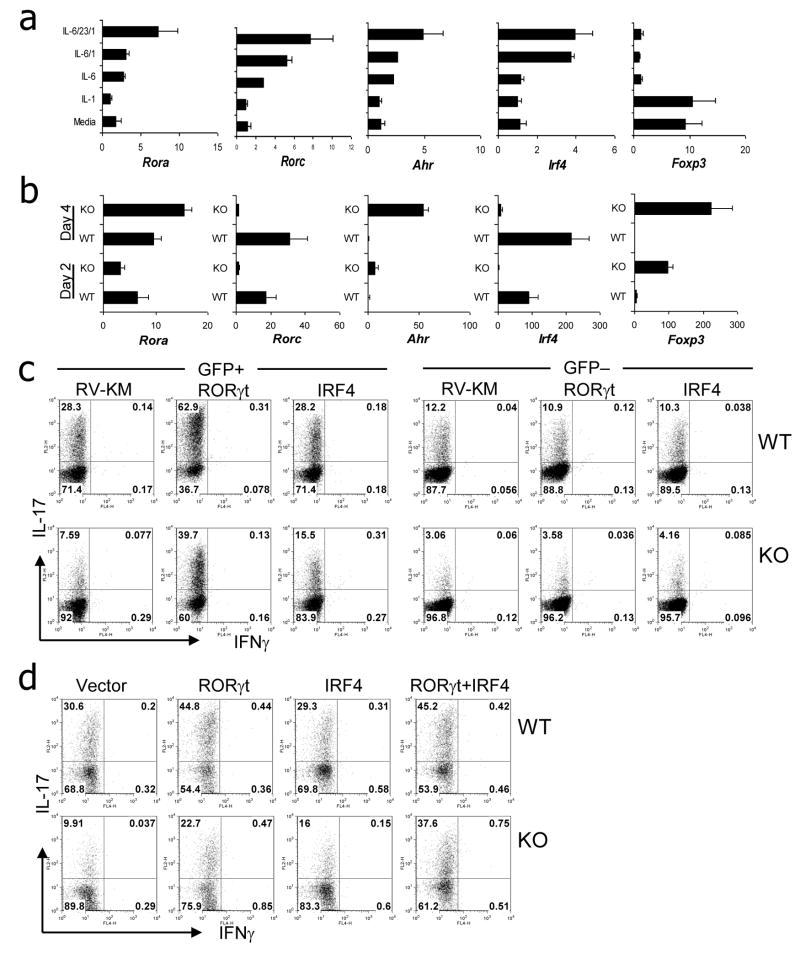
T helper (Th) 17 cells have been recently discovered in both mouse and human. Here we show that interleukin-1 (IL-1) signaling on T cells is critically required for the early programming of Th17 cell lineage and Th17 cell-mediated autoimmunity. IL-1 receptor1 expression in T cells, which was induced by IL-6, was necessary for the induction of experimental autoimmune encephalomyelitis and for early Th17 cell differentiation in vivo. Moreover, IL-1 signaling in T cells was required in dendritic cell-mediated Th17 cell differentiation from naive or regulatory precursors and IL-1 synergized with IL-6 and IL-23 to regulate Th17 cell differentiation and maintain cytokine expression in effector Th17 cells. Importantly, IL-1 regulated the expression of the transcription factors IRF4 and RORgammat during Th17 cell differentiation; overexpression of these two factors resulted in IL-1-independent Th17 cell polarization. Our data thus indicate a critical role of IL-1 in Th17 cell differentiation and this pathway may serve as a unique target for Th17 cell-mediated immunopathology.
Figures






Similar articles
-
The receptor SIGIRR suppresses Th17 cell proliferation via inhibition of the interleukin-1 receptor pathway and mTOR kinase activation.Immunity. 2010 Jan 29;32(1):54-66. doi: 10.1016/j.immuni.2009.12.003. Epub 2010 Jan 7. Immunity. 2010. PMID: 20060329 Free PMC article.
-
T helper 17 lineage differentiation is programmed by orphan nuclear receptors ROR alpha and ROR gamma.Immunity. 2008 Jan;28(1):29-39. doi: 10.1016/j.immuni.2007.11.016. Epub 2007 Dec 27. Immunity. 2008. PMID: 18164222 Free PMC article.
-
The protein tyrosine kinase Tec regulates a CD44highCD62L- Th17 subset.J Immunol. 2010 Nov 1;185(9):5111-9. doi: 10.4049/jimmunol.1001734. Epub 2010 Sep 24. J Immunol. 2010. PMID: 20870948
-
Signal transduction pathways and transcriptional regulation in the control of Th17 differentiation.Semin Immunol. 2007 Dec;19(6):400-8. doi: 10.1016/j.smim.2007.10.015. Epub 2007 Dec 31. Semin Immunol. 2007. PMID: 18166487 Free PMC article. Review.
-
Differentiation and function of pro-inflammatory Th17 cells.Microbes Infect. 2009 Apr;11(5):584-8. doi: 10.1016/j.micinf.2009.04.001. Epub 2009 Apr 14. Microbes Infect. 2009. PMID: 19371793 Free PMC article. Review.
Cited by
-
Epithelial-derived IL-18 regulates Th17 cell differentiation and Foxp3⁺ Treg cell function in the intestine.Mucosal Immunol. 2015 Nov;8(6):1226-36. doi: 10.1038/mi.2015.13. Epub 2015 Mar 4. Mucosal Immunol. 2015. PMID: 25736457 Free PMC article.
-
A critical role for Th17 cell-derived TGF-β1 in regulating the stability and pathogenicity of autoimmune Th17 cells.Exp Mol Med. 2021 May;53(5):993-1004. doi: 10.1038/s12276-021-00632-9. Epub 2021 May 28. Exp Mol Med. 2021. PMID: 34050263 Free PMC article.
-
Protein kinase CK2 governs the molecular decision between encephalitogenic TH17 cell and Treg cell development.Proc Natl Acad Sci U S A. 2016 Sep 6;113(36):10145-50. doi: 10.1073/pnas.1523869113. Epub 2016 Aug 23. Proc Natl Acad Sci U S A. 2016. PMID: 27555590 Free PMC article.
-
IL-17 mediates immunopathology in the absence of IL-10 following Leishmania major infection.PLoS Pathog. 2013 Mar;9(3):e1003243. doi: 10.1371/journal.ppat.1003243. Epub 2013 Mar 21. PLoS Pathog. 2013. PMID: 23555256 Free PMC article.
-
Tumor progression locus 2 differentially regulates IFNγ and IL-17 production by effector CD4+ T cells in a T cell transfer model of colitis.PLoS One. 2015 Mar 17;10(3):e0119885. doi: 10.1371/journal.pone.0119885. eCollection 2015. PLoS One. 2015. PMID: 25781948 Free PMC article.
References
-
- Acosta-Rodriguez EV, Napolitani G, Lanzavecchia A, Sallusto F. Interleukins 1beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat Immunol. 2007;8:942–949. - PubMed
-
- Atarashi K, Nishimura J, Shima T, Umesaki Y, Yamamoto M, Onoue M, Yagita H, Ishii N, Evans R, Honda K, Takeda K. ATP drives lamina propria T(H)17 cell differentiation. Nature. 2008;455:808–812. - PubMed
-
- Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–238. - PubMed
-
- Brustle A, Heink S, Huber M, Rosenplanter C, Stadelmann C, Yu P, Arpaia E, Mak TW, Kamradt T, Lohoff M. The development of inflammatory T(H)-17 cells requires interferon-regulatory factor 4. Nat Immunol. 2007;8:958–966. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
