

Quiescent T lymphocytes as an inducible virus reservoir in HIV-1 infection
- PMID: 1925601
- PMCID: PMC9524215
- DOI: 10.1126/science.1925601
Quiescent T lymphocytes as an inducible virus reservoir in HIV-1 infection
Abstract
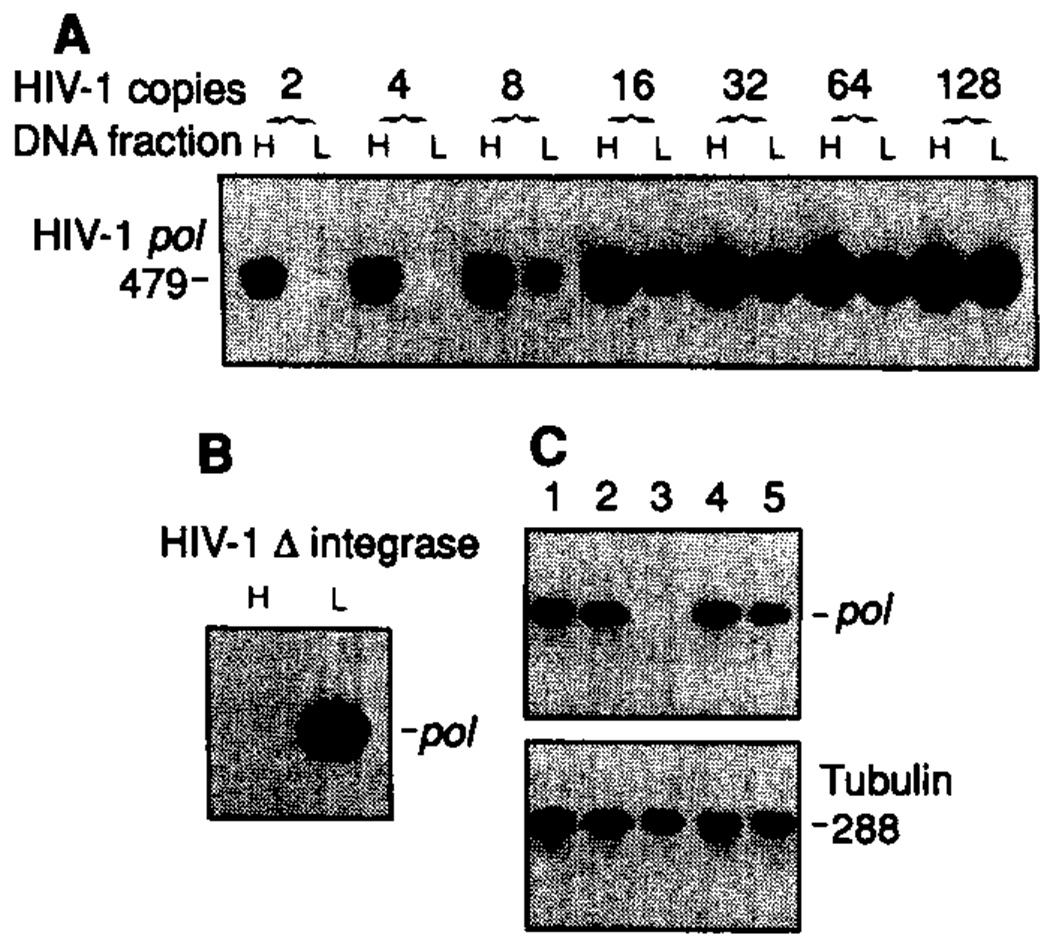
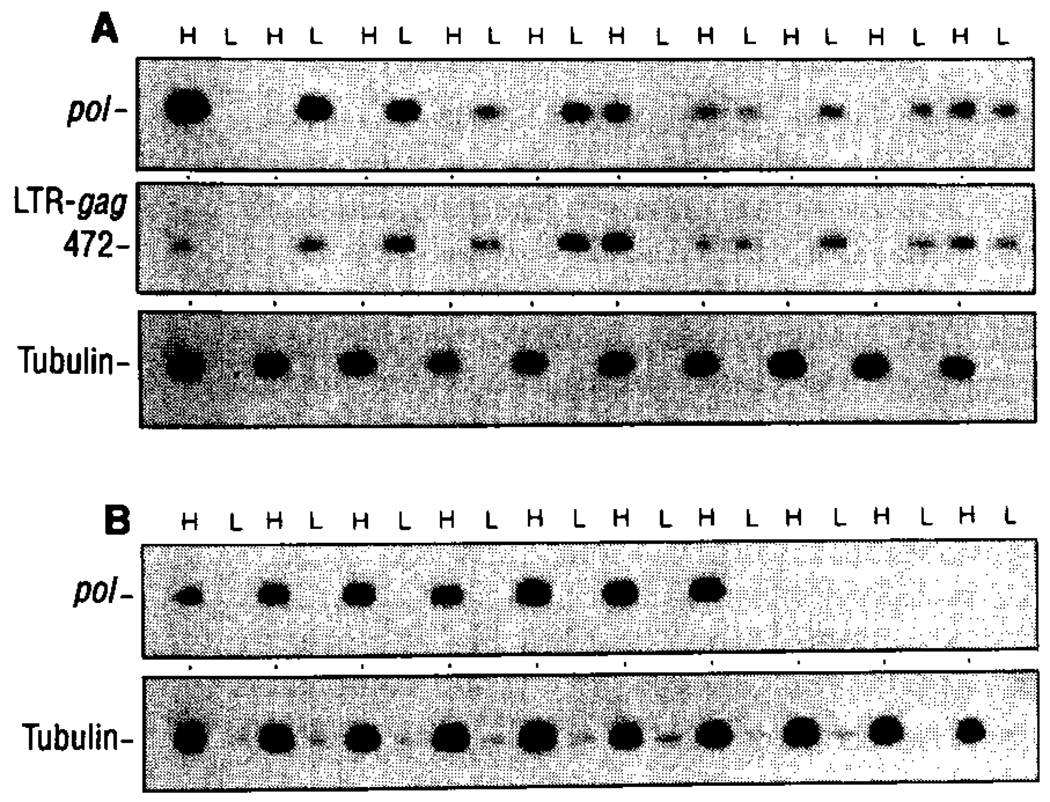
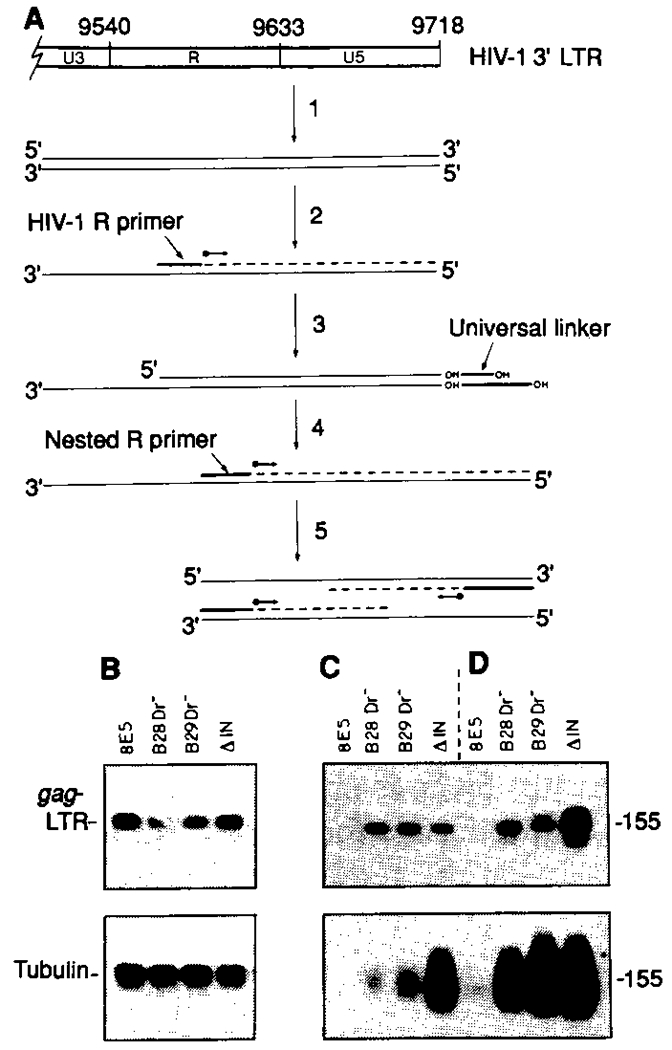
To better understand the basis for human immunodeficiency virus type 1 (HIV-1) persistence and latency, the form in which viral DNA exists in the peripheral T lymphocyte reservoir of infected individuals was investigated. In asymptomatic individuals, HIV-1 was harbored predominantly as full-length, unintegrated complementary DNA. These extrachromosomal DNA forms retained the ability to integrate upon T cell activation in vitro. In patients with acquired immunodeficiency syndrome (AIDS), there was an increase in integrated relative to extrachromosomal DNA forms. By analysis of DNA from patient lymphocyte subpopulations depleted of human lymphocyte antigen-Dr receptor-positive cells, quiescent T cells were identified as the source of extrachromosomal HIV-1 DNA. Thus quiescent T lymphocytes may be a major and inducible HIV-1 reservoir in infected individuals.
Figures
Similar articles
-
Establishment of a stable, inducible form of human immunodeficiency virus type 1 DNA in quiescent CD4 lymphocytes in vitro.J Virol. 1995 May;69(5):2977-88. doi: 10.1128/JVI.69.5.2977-2988.1995. J Virol. 1995. PMID: 7707524 Free PMC article.
-
Next-Generation Sequencing in a Direct Model of HIV Infection Reveals Important Parallels to and Differences from In Vivo Reservoir Dynamics.J Virol. 2020 Apr 16;94(9):e01900-19. doi: 10.1128/JVI.01900-19. Print 2020 Apr 16. J Virol. 2020. PMID: 32051279 Free PMC article.
-
HIV-1 replication is controlled at the level of T cell activation and proviral integration.EMBO J. 1990 May;9(5):1551-60. doi: 10.1002/j.1460-2075.1990.tb08274.x. EMBO J. 1990. PMID: 2184033 Free PMC article.
-
The role of integration and clonal expansion in HIV infection: live long and prosper.Retrovirology. 2018 Oct 23;15(1):71. doi: 10.1186/s12977-018-0448-8. Retrovirology. 2018. PMID: 30352600 Free PMC article. Review.
-
Low Inducibility of Latent Human Immunodeficiency Virus Type 1 Proviruses as a Major Barrier to Cure.J Infect Dis. 2021 Feb 15;223(12 Suppl 2):13-21. doi: 10.1093/infdis/jiaa649. J Infect Dis. 2021. PMID: 33586775 Free PMC article. Review.
Cited by
-
Biomimetic cell stimulation with a graphene oxide antigen-presenting platform for developing T cell-based therapies.Nat Nanotechnol. 2024 Dec;19(12):1914-1922. doi: 10.1038/s41565-024-01781-4. Epub 2024 Sep 23. Nat Nanotechnol. 2024. PMID: 39313679
-
Impact of fluctuation in frequency of human immunodeficiency virus/simian immunodeficiency virus reactivation during antiretroviral therapy interruption.Proc Biol Sci. 2020 Aug 26;287(1933):20200354. doi: 10.1098/rspb.2020.0354. Epub 2020 Aug 19. Proc Biol Sci. 2020. PMID: 32811309 Free PMC article.
-
Neisseria gonorrhoeae enhances HIV-1 infection of primary resting CD4+ T cells through TLR2 activation.J Immunol. 2010 Mar 15;184(6):2814-24. doi: 10.4049/jimmunol.0902125. Epub 2010 Feb 10. J Immunol. 2010. PMID: 20147631 Free PMC article.
-
An anti-CD45RO immunotoxin eliminates T cells latently infected with HIV-1 in vitro.Proc Natl Acad Sci U S A. 1999 Sep 28;96(20):11482-5. doi: 10.1073/pnas.96.20.11482. Proc Natl Acad Sci U S A. 1999. PMID: 10500202 Free PMC article.
-
Vpr is required for efficient Nef expression from unintegrated human immunodeficiency virus type 1 DNA.J Virol. 2007 Oct;81(19):10515-23. doi: 10.1128/JVI.00947-07. Epub 2007 Jul 25. J Virol. 2007. PMID: 17652391 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
