

doi: 10.1016/S0083-6729(08)00620-1.
Retinal insulin receptor signaling in hyperosmotic stress
Affiliations
- PMID: 19251051
- PMCID: PMC2670484
- DOI: 10.1016/S0083-6729(08)00620-1
Item in Clipboard
Retinal insulin receptor signaling in hyperosmotic stress
Vitam Horm.
2009.
Abstract
In the diabetic eye, the increased accumulation of sorbitol in the retina has been implicated in the pathogenesis of diabetic retinopathy (DR). Neurodegeneration is an important component of DR as demonstrated by increased neural apoptosis in the retina during experimental and human diabetes. Insulin receptor (IR) activation has been shown to rescue retinal neurons from apoptosis through a phosphoinositide 3-kinase and protein kinase B (Akt) survival cascade. In this study, we examined the IR signaling in sorbitol-induced hyperosmotic stressed retinas.
Figures

Concentration dependent Sorbitol-induced activation of p38 MAP kinase. Various concentrations of sorbitol treated retinal samples were subjected to Western blot analysis with anti-phospho-p38MAP kinase (A) and anti-p38 MAP kinase (B) antibodies.
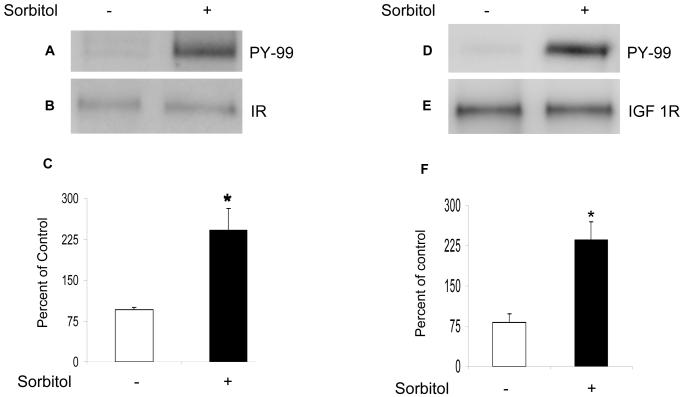
Sorbitol-induced activation of IR and IGF-1R. Retinal proteins from control and 1.0 M sorbitol-treated organotypic cultures were immunoprecipitated with anti-IRß antibody followed by Western blot analysis with anti-PY99 antibody (A). The blots were stripped and reprobed with anti-IRß antibody to ensure equal amounts of IR in each immunoprecipitate (B). Densities were calculated from the immunoblots and the results are expressed as percentage of PY99/IR. Absence of sorbitol was taken as 100 percent (C). Data mean ± SD, n=3, *p<0.001. Sorbitol-induced activation of IGF 1R. Retinal proteins from the control and 1.0 M sorbitol-treated organotypic cultures were immunoprecipitated with anti-IGF 1R antibody followed by Western blot analysis with anti-PY99 antibody (D). The blots were stripped and reprobed with anti-IGF 1R antibody to ensure equal amount of IGF 1R in each immunoprecipitate (E). Densities were calculated from the immunoblots and the results are expressed as percentage of PY99/IR. Absence of sorbitol was taken as 100 percent (F). Data mean ± SD, n=3, *p<0.001.
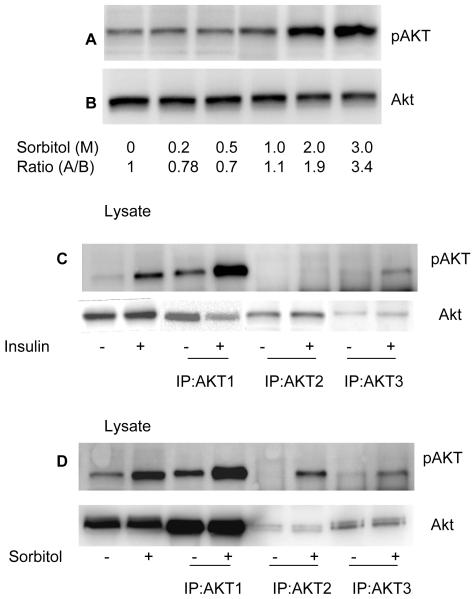
Sorbitol-induced activation of Akt isoforms. Various concentrations of sorbitol treated retinal samples were subjected to Western blot analysis with anti-pAkt (S473) (A) and anti-Akt (B) antibodies. Retinas were incubated in culture and treated with either insulin (1 μM) or sorbitol (3M). Retinas were lysed and the proteins were subjected to immunoprecipitation with anti-Akt1, anti-Akt2 and Anti-Akt3 antibodies followed by Western blot analysis with anti-pAkt (S473) (C and D) antibody. Input, 30 μg of retina lysates stimulated with either presence or absence of insulin or sorbitol.
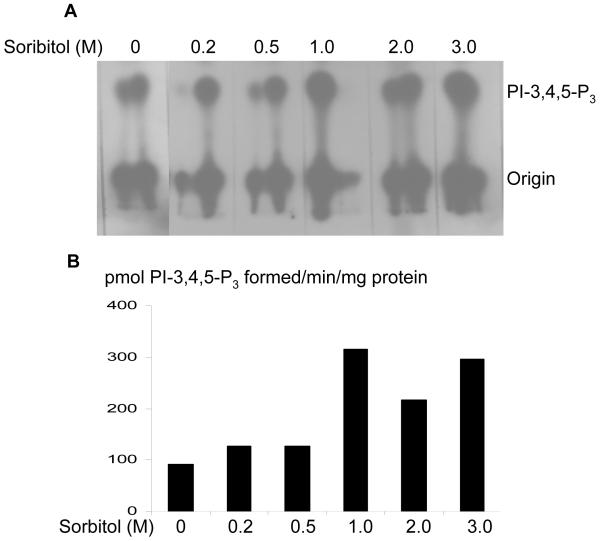
Sorbitol-induced activation insulin receptor associated PI3K activity. Retinas were cultured in DMEM and treated with various concentrations of sorbitol (0-3 M) for 30 min at 37 °C. TLC autoradiogram of PI3K activity measured in anti-IRβ immunoprecipitates of retinas using PI-4,5-P2 and [γ32P]ATP as substrates (A). The radioactive spots of PI-3,4,5-P3 were scraped from the TLC plate and counted (B).

PI3K-independent activation of Akt. Rat retinas were pre-incubated in DMEM medium with or without the PI3K inhibitor LY294002 (50 μM) for 30 min prior to either 1μM insulin or 3 M sorbitol. Thirty micrograms of retina lysate were subjected to Western blot analysis with anti-phospho-Akt (Ser 473) (A). The blot was stripped and reprobed with anti-Akt (B) and anti-actin (C) antibodies. PI3K activation is independent of p38 MAP kinase activation. Rat retinas were pre-incubated in DMEM medium with or without the PI3K inhibitor LY294002 (50 μM) or MAP kinase inhibitor SB203580 (50 μM) for 30 min prior to the treatment of either 1μM insulin or 3 M sorbitol. Thirty micrograms of retina lysates were subjected to Western blot analysis with anti-phospho-p38 MAP kinase(D). The blot was stripped and reprobed with anti-p38 MAP kinase (E) and anti-actin (F) antibodies.
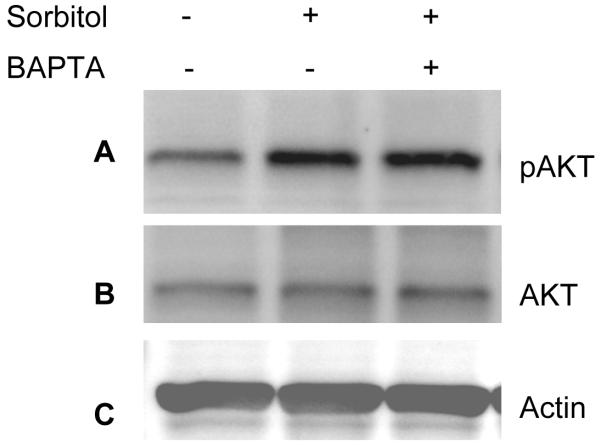
Sorbitol-induced activation of Akt is calcium-independent. Rat retinas were pre-incubated in DMEM medium with or without 15 μM BAPTA for 30 min prior to the treatment of 1 M sorbitol for additional 30 min. Thirty micrograms of retina lysates were subjected to Western blot analysis with anti-pAkt (S473) (A) anti-Akt (B), anti-actin (C) and anti-PY99 (D) antibodies.
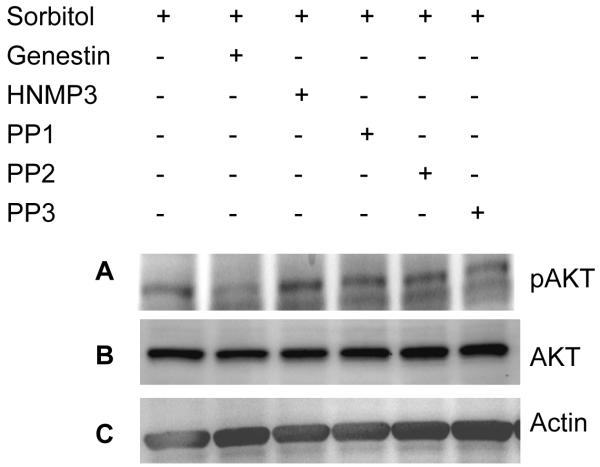
Tyrosine kinase induced-activation of Akt. Rat retinas were pre-incubated in DMEM medium with or without 100 μM genestin or HNMP3 or PP1 or PP2 or PP3 for 30 min prior to the treatment of 1 M sorbitol for 30 min. Thirty micrograms of retina lysates were subjected to Western blot analysis with anti-pAkt (S473) (A) anti-Akt (B) and anti-actin (C) antibodies.
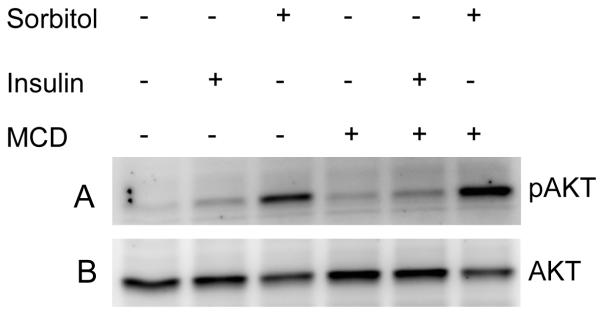
Cholesterol-depletion results in the activation of Akt. Rat retinas were incubated in DMEM medium with or without MCD prior to the treatment of 1 M sorbitol for 30 min. Thirty micrograms of retina lysates were subjected to Western blot analysis with anti-pAkt (S473) (A) and anti-Akt (B) antibodies.
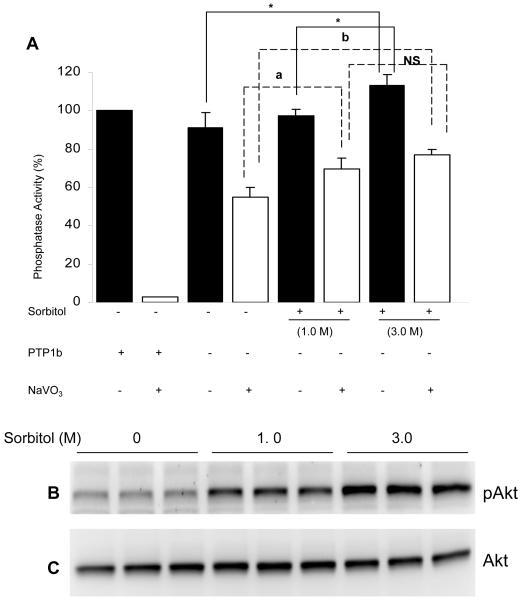
Sorbitol-induced activation of phosphatase activity. Retinal proteins from control and sorbitol-treated (1.0 or 3.0 M) or untreated organotypic cultures were subjected to either Western blotting analysis with anti-pAkt (B) and anti-Akt (C) antibodies or measured the phosphatase activity using pNPP as substrate (A). Open bars represent the activity in the presence of 1mM sodium vanadate. Purified GST-PTP1B (5μg) was used as positive control. Data are Data mean ± SD, n= three independent experiments 3, *p<0.001.
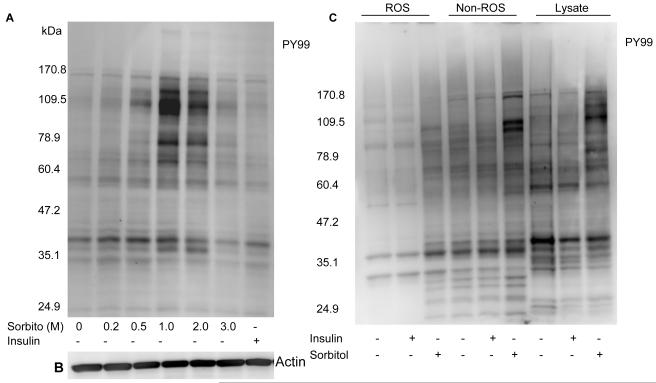
Sorbitol-induced tyrosine phosphorylation of several retinal proteins. Retinas were cultured in DMEM in the presence or the absence of either 1 μM insulin or various concentrations of sorbitol for 30 min at 37 °C. Thirty micrograms of retinal proteins were subjected to Western blot analysis with anti-PY 99 antibody (A). The blot was reprobed with the anti-actin antibody to ensure equal amount of protein in each lane (B). ROS and non-ROS proteins were subjected to Western blot analysis with the anti-PY99 antibody (C).
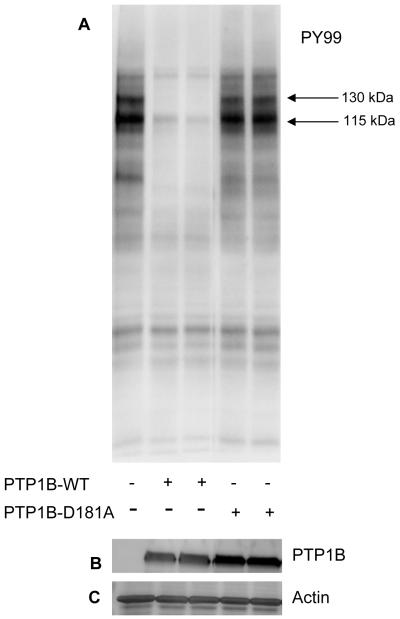
PTP1B dephosphorylates the sorbitol-induced tyrosine phosphorylated proteins in vitro. Sorbitol-treated retinal proteins were incubated with either wild type PTP1B or catalytically inactive PTP1B (D181A) for 15 min at 37 °C. At the end of the reaction, proteins were subjected to SDS-PAGE followed by Western blot analysis with anti-PY99 antibody (A). The blot was reprobed with anti-GST (B) and anti-actin (C) antibodies.

Interaction of Cbl with p85 subunit of PI3K. Sorbitol (1.0 M) treated or untreated retinal lysates were subjected to GST-pull down assay with GST-p85 full-length fusion protein followed by Western blot analysis with anti-Cbl antibody (A). Sorbitol treated and untreated retinal lysates were subjected to immunoprecipitation with anti-Cbl antibody followed by either Western blot analysis with anti-PY99 antibody (B) or measured the Cbl associated PI3K activity (C). Sorbitol treated and untreated retinal lysates were immunoprecipitated with anti-ubiquitin antibody followed by Western blot analysis with anti-p85 subunit of PI3K antibody (D).
Similar articles
-
PDGF- and insulin/IGF-1-specific distinct modes of class IA PI 3-kinase activation in normal rat retinas and RGC-5 retinal ganglion cells.Invest Ophthalmol Vis Sci. 2008 Aug;49(8):3687-98. doi: 10.1167/iovs.07-1455. Epub 2008 Apr 17. Invest Ophthalmol Vis Sci. 2008. PMID: 18421086
-
Insulin growth factor 1 receptor/PI3K/AKT survival pathway in outer segment membranes of rod photoreceptors.Invest Ophthalmol Vis Sci. 2008 Nov;49(11):4765-73. doi: 10.1167/iovs.08-2286. Epub 2008 Jun 19. Invest Ophthalmol Vis Sci. 2008. PMID: 18566464 Free PMC article.
-
Interaction of the retinal insulin receptor beta-subunit with the p85 subunit of phosphoinositide 3-kinase.Biochemistry. 2004 May 18;43(19):5637-50. doi: 10.1021/bi035913v. Biochemistry. 2004. PMID: 15134438
-
Characterization of insulin signaling in rat retina in vivo and ex vivo.Am J Physiol Endocrinol Metab. 2003 Oct;285(4):E763-74. doi: 10.1152/ajpendo.00507.2002. Epub 2003 Jun 10. Am J Physiol Endocrinol Metab. 2003. PMID: 12799319
-
Aldose reductase induced by hyperosmotic stress mediates cardiomyocyte apoptosis: differential effects of sorbitol and mannitol.J Biol Chem. 2003 Oct 3;278(40):38484-94. doi: 10.1074/jbc.M211824200. Epub 2003 Jul 24. J Biol Chem. 2003. PMID: 12881532
Cited by
-
Phosphoinositide 3-kinase signaling in the vertebrate retina.J Lipid Res. 2010 Jan;51(1):4-22. doi: 10.1194/jlr.R000232. J Lipid Res. 2010. PMID: 19638643 Free PMC article. Review.
-
Activation of sorbitol pathway in metabolic syndrome and increased susceptibility to cataract in Wistar-Obese rats.Mol Vis. 2012;18:495-503. Epub 2012 Feb 24. Mol Vis. 2012. PMID: 22393276 Free PMC article.
-
The role of insulin-like growth factor-I in the physiopathology of hearing.Front Mol Neurosci. 2011 Jul 25;4:11. doi: 10.3389/fnmol.2011.00011. eCollection 2011. Front Mol Neurosci. 2011. PMID: 21845174 Free PMC article.
-
Reducing Akt2 in retinal pigment epithelial cells causes a compensatory increase in Akt1 and attenuates diabetic retinopathy.Nat Commun. 2022 Oct 13;13(1):6045. doi: 10.1038/s41467-022-33773-0. Nat Commun. 2022. PMID: 36229454 Free PMC article.
-
Nrn1 Overexpression Attenuates Retinal Ganglion Cell Apoptosis, Promotes Axonal Regeneration, and Improves Visual Function Following Optic Nerve Crush in Rats.J Mol Neurosci. 2021 Jan;71(1):66-79. doi: 10.1007/s12031-020-01627-3. Epub 2020 Jun 30. J Mol Neurosci. 2021. PMID: 32607759
References
-
- Ahmed Z, Smith BJ, Pillay TS. The APS adapter protein couples the insulin receptor to the phosphorylation of c-Cbl and facilitates ligand-stimulated ubiquitination of the insulin receptor. FEBS Lett. 2000;475:31–34. - PubMed
-
- Asnaghi V, Gerhardinger C, Hoehn T, Adeboje A, Lorenzi M. A role for the polyol pathway in the early neuroretinal apoptosis and glial changes induced by diabetes in the rat. Diabetes. 2003;52:506–511. - PubMed
-
- Barber AJ, Nakamura M, Wolpert EB, Reiter CE, Seigel GM, Antonetti DA, Gardner TW. Insulin rescues retinal neurons from apoptosis by a phosphatidylinositol 3-kinase/Akt-mediated mechanism that reduces the activation of caspase-3. J Biol Chem. 2001;276:32814–32821. - PubMed
-
- Barnett PA, Gonzalez RG, Chylack LT, Jr., Cheng HM. The effect of oxidation on sorbitol pathway kinetics. Diabetes. 1986;35:426–432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
