

Review
doi: 10.1016/j.cell.2009.01.036.
Fidelity at the molecular level: lessons from protein synthesis
Affiliations
- PMID: 19239893
- PMCID: PMC3691815
- DOI: 10.1016/j.cell.2009.01.036
Item in Clipboard
Review
Fidelity at the molecular level: lessons from protein synthesis
Cell.
.
Abstract
The faithful and rapid translation of genetic information into peptide sequences is an indispensable property of the ribosome. The mechanistic understanding of strategies used by the ribosome to achieve both speed and fidelity during translation results from nearly a half century of biochemical and structural studies. Emerging from these studies is the common theme that the ribosome uses local as well as remote conformational switches to govern induced-fit mechanisms that ensure accuracy in codon recognition during both tRNA selection and translation termination.
Figures
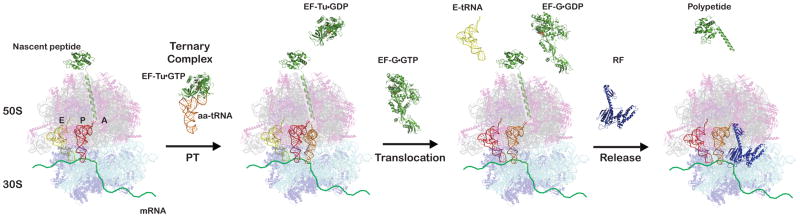
During the elongation cycle, peptidyl transfer (PT) takes place in which a ternary complex composed of the elongation factor EF-Tu, aminoacyl-tRNA (aa-tRNA), and GTP is deposited into the aminoacyl (A) site and reacts with the peptidyl-tRNA, elongating the nascent peptide by one amino acid. Subsequent translocation of the mRNA in the ribosome is mediated by elongation factor EF-G, which couples the energy of GTP hydrolysis to directional movement of the mRNA-tRNA complex. As a result, the peptidyl-tRNA and the deacylated tRNA move from the A and peptidyl (P) sites into the P and exit (E) sites, respectively. Termination of protein synthesis occurs when a stop codon enters the A site. Stop codons are recognized by class I release factors (RFs), which trigger a hydrolytic reaction that results in the release of the growing polypeptide chain from the tRNA. Molecular figures shown here were constructed and modified from PDB files (3D5A, 1GIX, 1COS, 1DAR, 1TTT, 1TUI, 2BV3 and 2VB1) using Pymol (DeLano Scientific) to depict the desired portions of the translation cycle. The E-site tRNA was omitted from the crystal structure in panel 2 and faded in panel 4 and to indicate the ambiguity of its status.
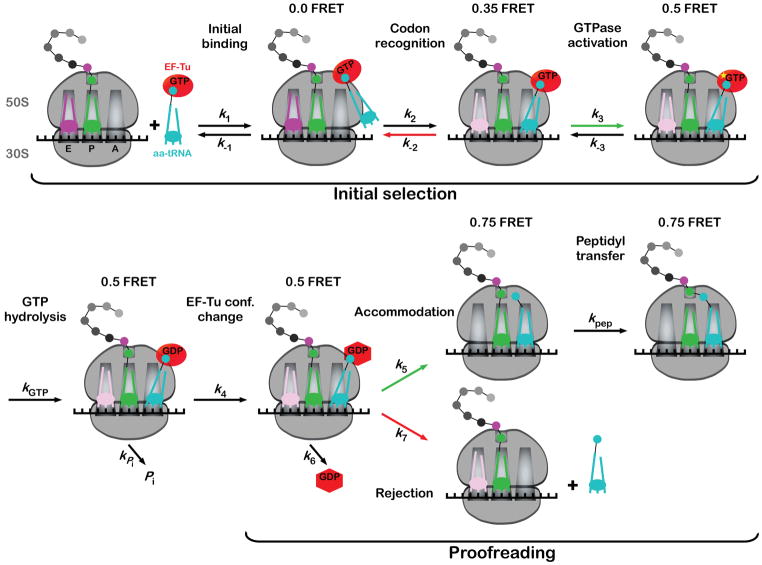
The view follows a tRNA through the selection process with defined steps indicated. The scheme is largely based on accumulated pre-steady state kinetic data (reviewed in Rodnina et al., 2005) from experiments performed in the absence of E-site tRNA. Predicted correlated FRET values are indicated at the top of each intermediate (Blanchard et al., 2004a). Green arrows indicate rates that are accelerated for cognate tRNA, whereas red arrows depict rates that are higher for near-cognate tRNA. We note that the occupancy and the role of the E-site tRNA following the codon-recognition intermediate is controversial and as such, the E-site tRNA is shown in a lighter color subsequent to this stage.
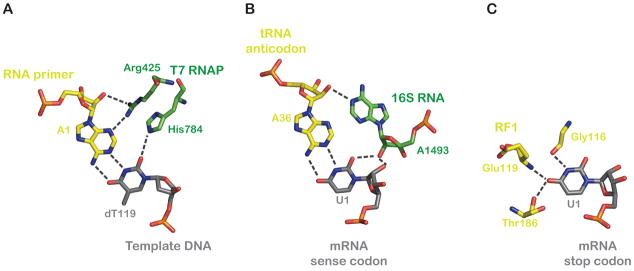
(Left) T7 RNAP (green) recognizes correct base-pairs between RNA (yellow) and DNA (grey) by a “molecular-caliper” mechanism, wherein side groups of specific residues measure the invariant properties of the Watson-Crick base-pair geometry (PDB 1MSW) (Yin and Steitz, 2002). (Center) The type-I A minor interaction at the first base-pair between the codon and anticodon (PDB 2J00) (Selmer et al., 2006). (Right) Recognition of U1 of the UAA stop codon by release factor 1 (RF1) (PDB 3D5A) (Laurberg et al., 2008). All molecular representations were generated using PyMol (DeLano Scientific).
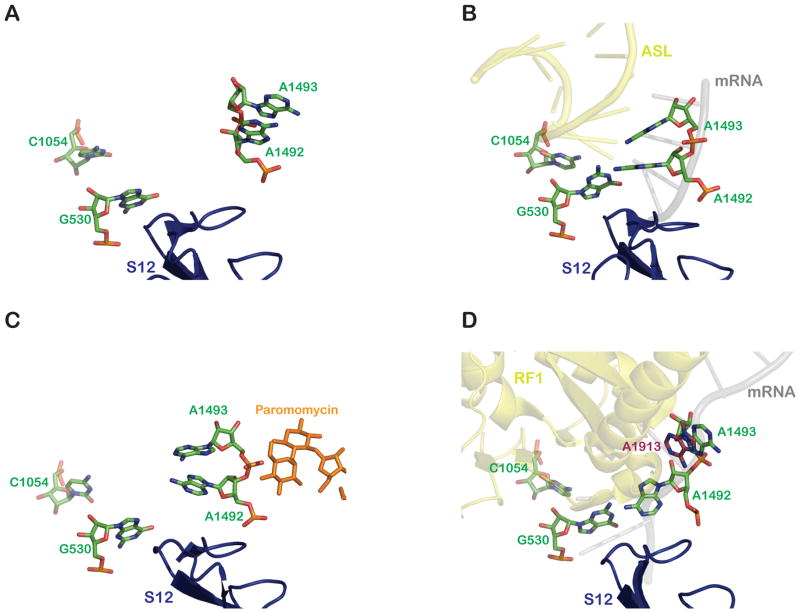
A comparison of the relative positions of key decoding center nucleotides (green) in an A site-vacant 30S ribosome subunit (A) (PDB 1J5E) (Wimberly et al., 2000), (B) in an ASL (yellow) -bound 30S subunit (PDB 1IBM) (Ogle et al., 2001), (C) in the presence of paromomycin (orange) (PDB 1IBK) (Ogle et al., 2001), and (D) in an RF1 (yellow) -bound 70S ribosome (PDB 3D5A) (Laurberg et al., 2008). 23S rRNA A1913: brick red. The movements of the nucleotides is described in details in the text. All molecular representations were generated using PyMol (DeLano Scientific).
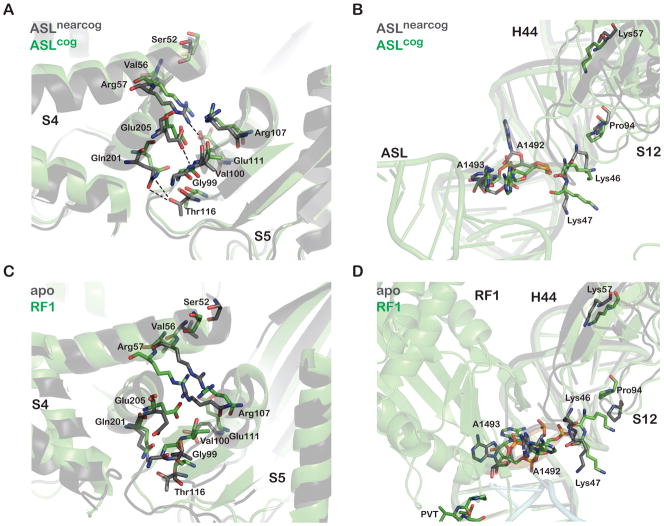
A. The S4/S5 small subunit proteins interface of cognate anticodon stemloop (ASL)-bound 30S ribosome subunit (PDB 1IBM) superimposed on the S4/S5 interface of near-cognate ASL-bound 30S ribosome subunit (PDB 1N34) (Ogle et al., 2002). S4 and S5 move apart as a result of cognate-ASL binding, breaking salt-bridges that favor their interaction. B. Superimposed structures as in A, now showing the relative positions of small subunit protein S12 and helix 44 of 16S RNA, where closer interactions are seen for the cognate structure. C. The S4/S5 interface of apo (PDB 2OW8) (Korostelev et al., 2006) superimposed on that of RF1-bound 70S ribosomes (PDB 3D5A) (Laurberg et al., 2008). In this case, distinct, more lateral movements are observed as a result of RF1 binding. D. Superimposed structures as in C, now showing the relative positions of S12 and helix 44, where in the context of RF1 binding, S12 moves away from helix 44.
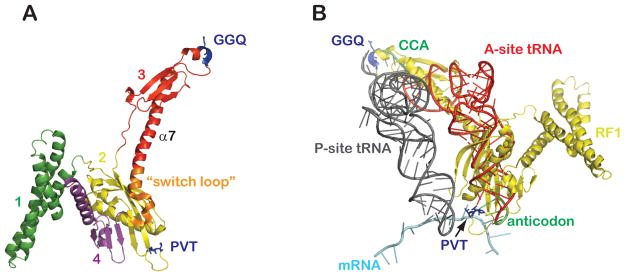
(A) Structure of release factor 1 (RF1) as bound to the 70S ribosome (PDB 3D5A) (Laurberg et al., 2008). Domains of RF1 are colored and numbered as indicated with the functional motifs highlighted in blue. (B) Ribosome-bound A-site tRNA (PDB 1GIX) (Yusupov et al., 2001) superimposed onto the structure of the same 70S termination complex (where P-site tRNA, mRNA, and RF1 are shown).
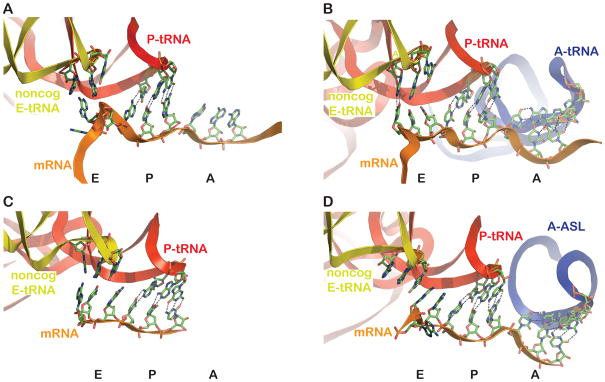
A. Structure of initiation-like complex, with a Shine-Dalgarno sequence upstream, from Jenner et al. (2007) (PDB 2HGR) containing non-cognate E-site tRNA, cognate P-site tRNA, and an empty A site. The mRNA adopts a conformation not appearing compatible with codon-anticodon interaction in the E site. B. Structure of elongation-like complex from same study as A (PDB 2HGP), also with non-cognate tRNA in the E site, and cognate P- and A- sites tRNAs. Continuous electron density, from the X-ray diffraction data, is observed between position 1 of the E site codon and the corresponding position of the E-site tRNA anticodon, as indicated with the dashed lines. C. Structure from Korostelev et al. (2006) (PDB 2OW8) with non-cognate E-site tRNA, cognate P-site tRNA and an empty A site (and no Shine-Dalgarno sequence). The mRNA adopts a conformation that appears to be compatible with codon-anticodon interactions, though none are documented with the non-cognate tRNA species bound. D. Structure from Selmer et al. (2006) (PDB 2J00) with ligands as in C, except that the A site is occupied with a cognate anticodon stemloop (ASL). The mRNA (most notably the first position of the E-site codon) and the E-site tRNA positions are distinct from those in C.
Similar articles
-
Quality control of mRNA decoding on the bacterial ribosome.Adv Protein Chem Struct Biol. 2012;86:95-128. doi: 10.1016/B978-0-12-386497-0.00003-7. Adv Protein Chem Struct Biol. 2012. PMID: 22243582 Review.
-
Polyamines accelerate codon recognition by transfer RNAs on the ribosome.Biochemistry. 2010 Aug 24;49(33):7179-89. doi: 10.1021/bi1009776. Biochemistry. 2010. PMID: 20666453 Free PMC article.
-
Quality control by the ribosome following peptide bond formation.Nature. 2009 Jan 8;457(7226):161-6. doi: 10.1038/nature07582. Epub 2008 Dec 17. Nature. 2009. PMID: 19092806 Free PMC article.
-
Altered tRNA dynamics during translocation on slippery mRNA as determinant of spontaneous ribosome frameshifting.Nat Commun. 2022 Jul 22;13(1):4231. doi: 10.1038/s41467-022-31852-w. Nat Commun. 2022. PMID: 35869111 Free PMC article.
-
[tRNA-binding centers of Escherichia coli ribosomes and their structural organization].Mol Biol (Mosk). 1984 Sep-Oct;18(5):1194-207. Mol Biol (Mosk). 1984. PMID: 6209546 Review. Russian.
Cited by
-
Knockout of AtMKK1 enhances salt tolerance and modifies metabolic activities in Arabidopsis.Plant Signal Behav. 2013 May;8(5):e24206. doi: 10.4161/psb.24206. Epub 2013 Mar 19. Plant Signal Behav. 2013. PMID: 23511202 Free PMC article.
-
Yeast Trm7 interacts with distinct proteins for critical modifications of the tRNAPhe anticodon loop.RNA. 2012 Oct;18(10):1921-33. doi: 10.1261/rna.035287.112. Epub 2012 Aug 21. RNA. 2012. PMID: 22912484 Free PMC article.
-
Estimating selection on synonymous codon usage from noisy experimental data.Mol Biol Evol. 2013 Jun;30(6):1438-53. doi: 10.1093/molbev/mst051. Epub 2013 Mar 14. Mol Biol Evol. 2013. PMID: 23493257 Free PMC article.
-
Insights into the base-pairing preferences of 8-oxoguanosine on the ribosome.Nucleic Acids Res. 2019 Oct 10;47(18):9857-9870. doi: 10.1093/nar/gkz701. Nucleic Acids Res. 2019. PMID: 31400119 Free PMC article.
-
Complex molecular dynamics in the spotlight.Nat Biotechnol. 2010 Jun;28(6):564-5. doi: 10.1038/nbt0610-564. Nat Biotechnol. 2010. PMID: 20531335 No abstract available.
References
-
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science. 2000;289:905–920. - PubMed
-
- Bilgin N, Claesens F, Pahverk H, Ehrenberg M. Kinetic properties of Escherichia coli ribosomes with altered forms of S12. J Mol Biol. 1992;224:1011–1027. - PubMed
-
- Blanchard SC, Gonzalez RL, Kim HD, Chu S, Puglisi JD. tRNA selection and kinetic proofreading in translation. Nat Struct Mol Biol. 2004a;11:1008–1014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
