

Conditional embryonic lethality to improve the sterile insect technique in Ceratitis capitata (Diptera: Tephritidae)
- PMID: 19173707
- PMCID: PMC2662800
- DOI: 10.1186/1741-7007-7-4
Conditional embryonic lethality to improve the sterile insect technique in Ceratitis capitata (Diptera: Tephritidae)
Abstract
Background: The sterile insect technique (SIT) is an environment-friendly method used in area-wide pest management of the Mediterranean fruit fly Ceratitis capitata (Wiedemann; Diptera: Tephritidae). Ionizing radiation used to generate reproductive sterility in the mass-reared populations before release leads to reduction of competitiveness.
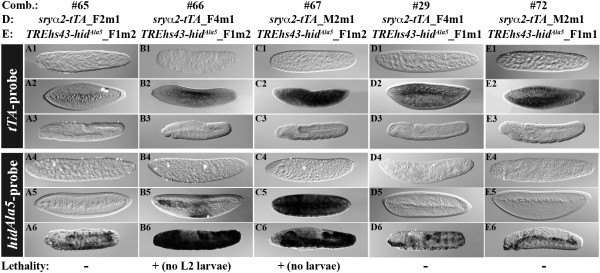
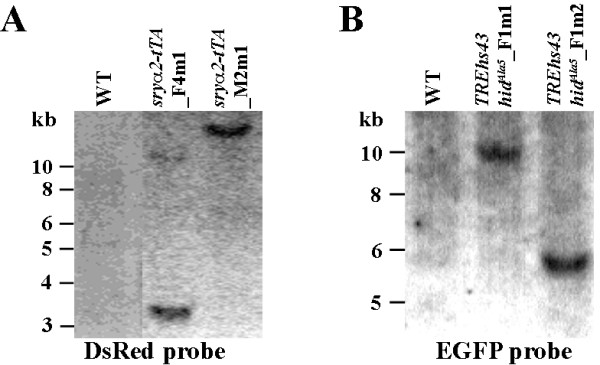
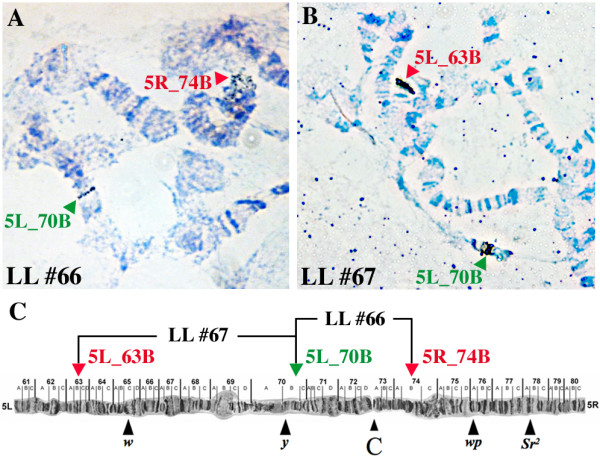
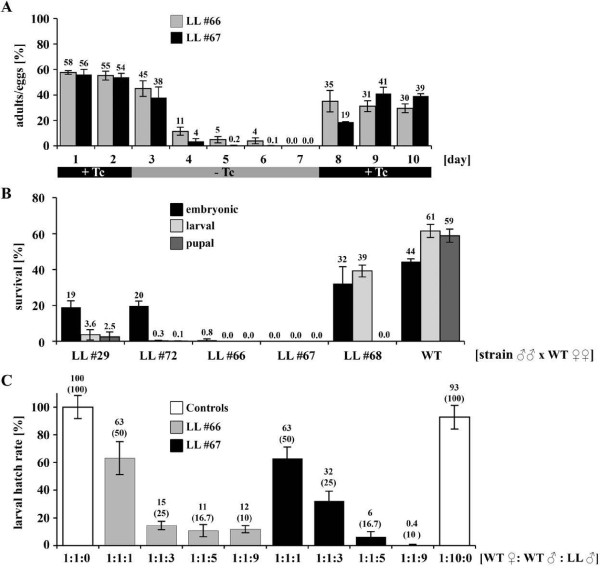
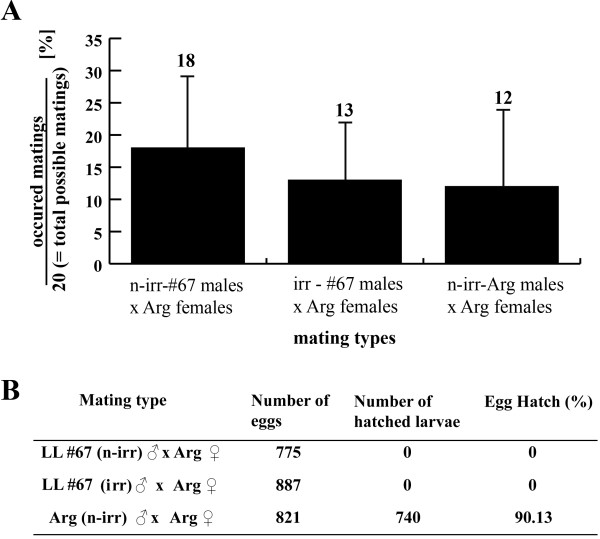
Results: Here, we present a first alternative reproductive sterility system for medfly based on transgenic embryonic lethality. This system is dependent on newly isolated medfly promoter/enhancer elements of cellularization-specifically-expressed genes. These elements act differently in expression strength and their ability to drive lethal effector gene activation. Moreover, position effects strongly influence the efficiency of the system. Out of 60 combinations of driver and effector construct integrations, several lines resulted in larval and pupal lethality with one line showing complete embryonic lethality. This line was highly competitive to wildtype medfly in laboratory and field cage tests.
Conclusion: The high competitiveness of the transgenic lines and the achieved 100% embryonic lethality causing reproductive sterility without the need of irradiation can improve the efficacy of operational medfly SIT programs.
Figures


Similar articles
-
Transgenic sexing system for Ceratitis capitata (Diptera: Tephritidae) based on female-specific embryonic lethality.Insect Biochem Mol Biol. 2013 Jan;43(1):1-8. doi: 10.1016/j.ibmb.2012.10.010. Epub 2012 Nov 5. Insect Biochem Mol Biol. 2013. PMID: 23137881
-
Genetic elimination of field-cage populations of Mediterranean fruit flies.Proc Biol Sci. 2014 Oct 7;281(1792):20141372. doi: 10.1098/rspb.2014.1372. Proc Biol Sci. 2014. PMID: 25122230 Free PMC article.
-
Delivery of Nucleic Acids through Embryo Microinjection in the Worldwide Agricultural Pest Insect, Ceratitis capitata.J Vis Exp. 2016 Oct 1;(116):54528. doi: 10.3791/54528. J Vis Exp. 2016. PMID: 27768087 Free PMC article.
-
How functional genomics will impact fruit fly pest control: the example of the Mediterranean fruit fly, Ceratitis capitata.BMC Genet. 2014;15 Suppl 2(Suppl 2):S11. doi: 10.1186/1471-2156-15-S2-S11. Epub 2014 Dec 1. BMC Genet. 2014. PMID: 25471105 Free PMC article. Review.
-
The Sterile Insect Technique: Success and Perspectives in the Neotropics.Neotrop Entomol. 2021 Apr;50(2):172-185. doi: 10.1007/s13744-020-00817-3. Epub 2020 Oct 28. Neotrop Entomol. 2021. PMID: 33113111 Review.
Cited by
-
Nanopore long-read RNA-seq and absolute quantification delineate transcription dynamics in early embryo development of an insect pest.Sci Rep. 2021 Apr 12;11(1):7878. doi: 10.1038/s41598-021-86753-7. Sci Rep. 2021. PMID: 33846393 Free PMC article.
-
Improvement and use of CRISPR/Cas9 to engineer a sperm-marking strain for the invasive fruit pest Drosophila suzukii.BMC Biotechnol. 2019 Dec 5;19(1):85. doi: 10.1186/s12896-019-0588-5. BMC Biotechnol. 2019. PMID: 31805916 Free PMC article.
-
Identification and characterization of four Drosophila suzukii cellularization genes and their promoters.BMC Genet. 2020 Dec 18;21(Suppl 2):146. doi: 10.1186/s12863-020-00939-y. BMC Genet. 2020. PMID: 33339500 Free PMC article.
-
Functionality of the GAL4/UAS system in Tribolium requires the use of endogenous core promoters.BMC Dev Biol. 2010 May 19;10:53. doi: 10.1186/1471-213X-10-53. BMC Dev Biol. 2010. PMID: 20482875 Free PMC article.
-
Molecular tools to create new strains for mosquito sexing and vector control.Parasit Vectors. 2018 Dec 24;11(Suppl 2):645. doi: 10.1186/s13071-018-3209-6. Parasit Vectors. 2018. PMID: 30583736 Free PMC article. Review.
References
-
- White IM, Elson-Harris MM. Fruit flies of economic significance: their identification and bionomics. Wallingford, Oxon, UK: CAB International; 1992.
-
- Enkerlin WR. Impact of fruit fly control programmes using the sterile insect technique. In: Dyck VA, Hendrichs J, Robinson AS, editor. Sterile Insect Technique – Principles and Practice in Area-wide Integrated Pest Management. Dordrecht, Netherlands: Springer; 2005. pp. 651–676.
-
- Knipling EF. Possibilities of insect control or eradication through the use of sexually sterile males. J Econ Entomol. 1955;48:459–462.
-
- Parker A, Mehta K. Sterile insect technique: a model for dose optimization for improved sterile insect quality. Fla Entomol. 2007;90:88–95.
-
- Hendrichs J, Robinson AS, Cayol JP, Enkerlin W. Medfly areawide sterile insect technique programmes for prevention, suppression or eradication: the importance of mating behavior studies. Fla Entomol. 2002;85:1–13.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
