

A gene expression signature shared by human mature oocytes and embryonic stem cells
- PMID: 19128516
- PMCID: PMC2628676
- DOI: 10.1186/1471-2164-10-10
A gene expression signature shared by human mature oocytes and embryonic stem cells
Abstract
Background: The first week of human pre-embryo development is characterized by the induction of totipotency and then pluripotency. The understanding of this delicate process will have far reaching implication for in vitro fertilization and regenerative medicine. Human mature MII oocytes and embryonic stem (ES) cells are both able to achieve the feat of cell reprogramming towards pluripotency, either by somatic cell nuclear transfer or by cell fusion, respectively. Comparison of the transcriptome of these two cell types may highlight genes that are involved in pluripotency initiation.
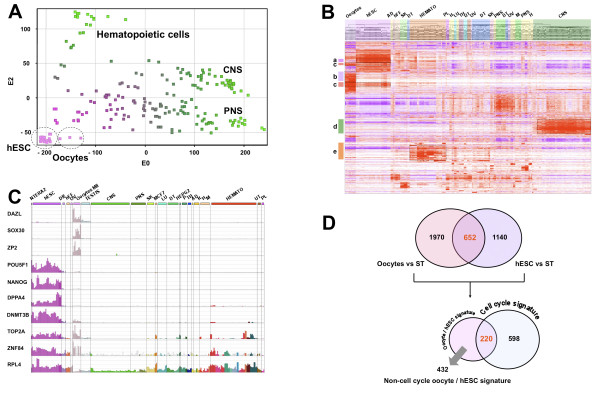
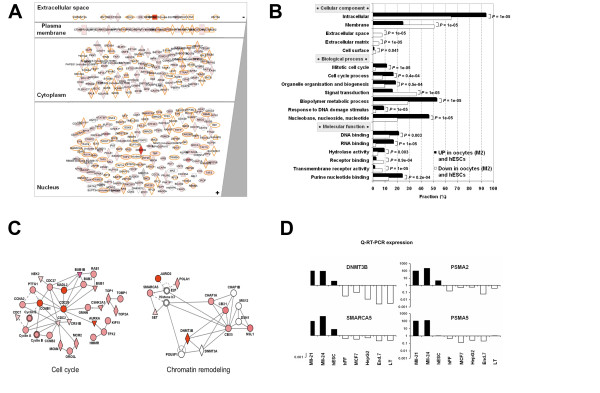
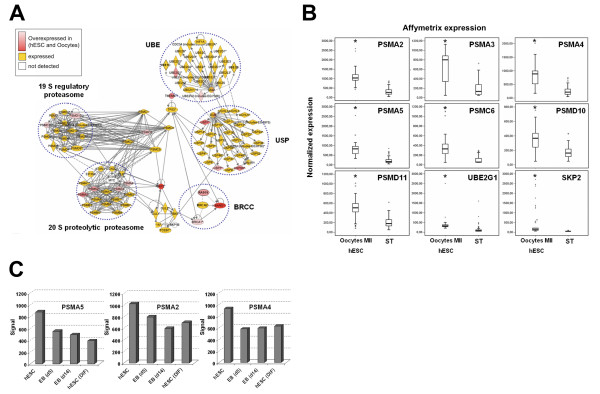
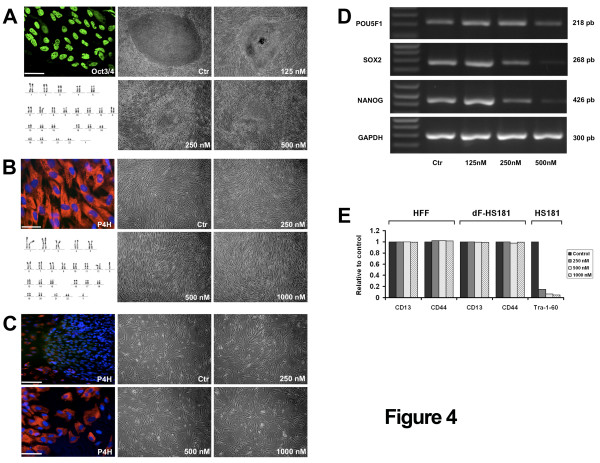
Results: Based on a microarray compendium of 205 samples, we compared the gene expression profile of mature MII oocytes and human ES cells (hESC) to that of somatic tissues. We identified a common oocyte/hESC gene expression profile, which included a strong cell cycle signature, genes associated with pluripotency such as LIN28 and TDGF1, a large chromatin remodelling network (TOP2A, DNMT3B, JARID2, SMARCA5, CBX1, CBX5), 18 different zinc finger transcription factors, including ZNF84, and several still poorly annotated genes such as KLHL7, MRS2, or the Selenophosphate synthetase 1 (SEPHS1). Interestingly, a large set of genes was also found to code for proteins involved in the ubiquitination and proteasome pathway. Upon hESC differentiation into embryoid bodies, the transcription of this pathway declined. In vitro, we observed a selective sensitivity of hESC to the inhibition of the activity of the proteasome.
Conclusion: These results shed light on the gene networks that are concurrently overexpressed by the two human cell types with somatic cell reprogramming properties.
Figures
Similar articles
-
Predicting distinct organization of transcription factor binding sites on the promoter regions: a new genome-based approach to expand human embryonic stem cell regulatory network.Gene. 2013 Dec 1;531(2):212-9. doi: 10.1016/j.gene.2013.09.011. Epub 2013 Sep 13. Gene. 2013. PMID: 24042128
-
Evaluation in mammalian oocytes of gene transcripts linked to epigenetic reprogramming.Reproduction. 2007 Oct;134(4):549-58. doi: 10.1530/REP-06-0315. Reproduction. 2007. PMID: 17890290
-
Characterization and gene expression profiling of five human embryonic stem cell lines derived in Taiwan.Methods Mol Biol. 2012;873:127-49. doi: 10.1007/978-1-61779-794-1_8. Methods Mol Biol. 2012. PMID: 22528352
-
Dynamic changes in gene expression during human early embryo development: from fundamental aspects to clinical applications.Hum Reprod Update. 2011 Mar-Apr;17(2):272-90. doi: 10.1093/humupd/dmq036. Epub 2010 Aug 17. Hum Reprod Update. 2011. PMID: 20716614 Free PMC article. Review.
-
A review of gene expression profiling of human embryonic stem cell lines and their differentiated progeny.Curr Stem Cell Res Ther. 2009 May;4(2):98-106. doi: 10.2174/157488809788167409. Curr Stem Cell Res Ther. 2009. PMID: 19442194 Review.
Cited by
-
Endothelial Jarid2/Jumonji is required for normal cardiac development and proper Notch1 expression.J Biol Chem. 2011 May 13;286(19):17193-204. doi: 10.1074/jbc.M110.205146. Epub 2011 Mar 14. J Biol Chem. 2011. PMID: 21402699 Free PMC article.
-
Structural asymmetry in the magnesium channel CorA points to sequential allosteric regulation.Proc Natl Acad Sci U S A. 2012 Nov 13;109(46):18809-14. doi: 10.1073/pnas.1209018109. Epub 2012 Oct 29. Proc Natl Acad Sci U S A. 2012. PMID: 23112165 Free PMC article.
-
ATP dependent chromatin remodeling enzymes in embryonic stem cells.Stem Cell Rev Rep. 2010 Mar;6(1):62-73. doi: 10.1007/s12015-010-9120-y. Stem Cell Rev Rep. 2010. PMID: 20148317 Free PMC article. Review.
-
Ontological aspects of pluripotency and stemness gene expression pattern in the rhesus monkey.Gene Expr Patterns. 2011 Mar-Apr;11(3-4):285-98. doi: 10.1016/j.gep.2011.02.001. Epub 2011 Feb 15. Gene Expr Patterns. 2011. PMID: 21329766 Free PMC article.
-
Essential role of maternal UCHL1 and UCHL3 in fertilization and preimplantation embryo development.J Cell Physiol. 2012 Apr;227(4):1592-603. doi: 10.1002/jcp.22876. J Cell Physiol. 2012. PMID: 21678411 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
