

Functional characterization of the vaccinia virus I5 protein
- PMID: 19077320
- PMCID: PMC2621143
- DOI: 10.1186/1743-422X-5-148
Functional characterization of the vaccinia virus I5 protein
Abstract
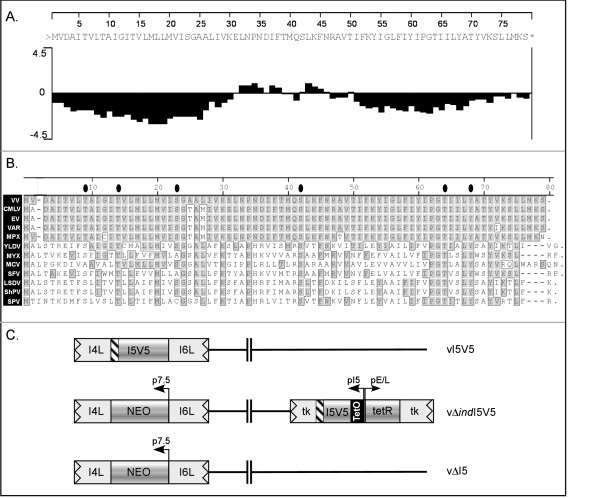
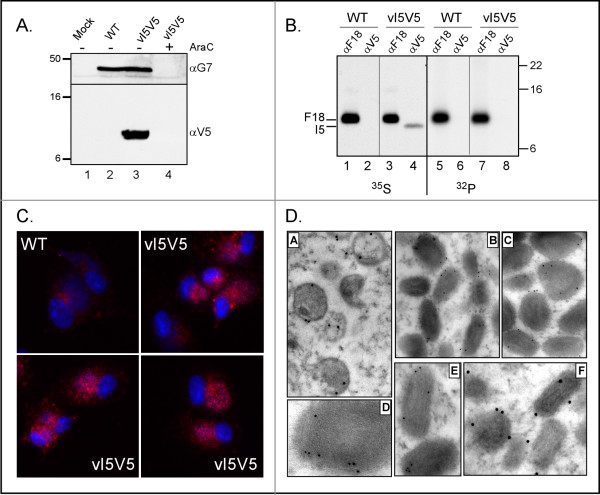
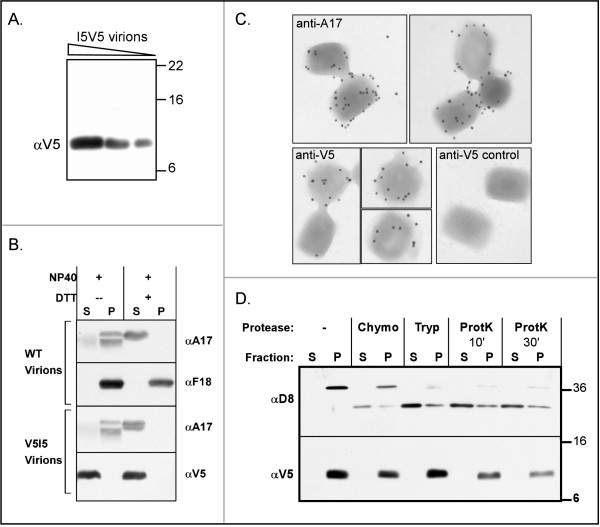
The I5L gene is one of approximately 90 genes that are conserved throughout the chordopoxvirus family, and hence are presumed to play vital roles in the poxvirus life cycle. Previous work had indicated that the VP13 protein, a component of the virion membrane, was encoded by the I5L gene, but no additional studies had been reported. Using a recombinant virus that encodes an I5 protein fused to a V5 epitope tag at the endogenous locus (vI5V5), we show here that the I5 protein is expressed as a post-replicative gene and that the approximately 9 kDa protein does not appear to be phosphorylated in vivo. I5 does not appear to traffic to any cellular organelle, but ultrastructural and biochemical analyses indicate that I5 is associated with the membranous components of assembling and mature virions. Intact virions can be labeled with anti-V5 antibody as assessed by immunoelectron microscopy, indicating that the C' terminus of the protein is exposed on the virion surface. Using a recombinant virus which encodes only a TET-regulated copy of the I5V5 gene (vDeltaindI5V5), or one in which the I5 locus has been deleted (vDeltaI5), we also show that I5 is dispensable for replication in tissue culture. Neither plaque size nor the viral yield produced in BSC40 cells or primary human fibroblasts are affected by the absence of I5 expression.
Figures

Similar articles
-
Vaccinia virus encodes I5, a small hydrophobic virion membrane protein that enhances replication and virulence in mice.J Virol. 2008 Oct;82(20):10071-8. doi: 10.1128/JVI.01355-08. Epub 2008 Aug 13. J Virol. 2008. PMID: 18701595 Free PMC article.
-
Vaccinia virus A28L gene encodes an essential protein component of the virion membrane with intramolecular disulfide bonds formed by the viral cytoplasmic redox pathway.J Virol. 2004 Mar;78(5):2348-56. doi: 10.1128/jvi.78.5.2348-2356.2004. J Virol. 2004. PMID: 14963131 Free PMC article.
-
Direct formation of vaccinia virus membranes from the endoplasmic reticulum in the absence of the newly characterized L2-interacting protein A30.5.J Virol. 2013 Nov;87(22):12313-26. doi: 10.1128/JVI.02137-13. Epub 2013 Sep 11. J Virol. 2013. PMID: 24027302 Free PMC article.
-
Vaccinia virus morphogenesis and dissemination.Trends Microbiol. 2008 Oct;16(10):472-9. doi: 10.1016/j.tim.2008.07.009. Epub 2008 Sep 12. Trends Microbiol. 2008. PMID: 18789694 Review.
-
In a nutshell: structure and assembly of the vaccinia virion.Adv Virus Res. 2006;66:31-124. doi: 10.1016/S0065-3527(06)66002-8. Adv Virus Res. 2006. PMID: 16877059 Review.
Cited by
-
Biogenesis of the vaccinia virus membrane: genetic and ultrastructural analysis of the contributions of the A14 and A17 proteins.J Virol. 2013 Jan;87(2):1083-97. doi: 10.1128/JVI.02529-12. Epub 2012 Nov 7. J Virol. 2013. PMID: 23135725 Free PMC article.
-
Viral miniproteins.Annu Rev Microbiol. 2014;68:21-43. doi: 10.1146/annurev-micro-091313-103727. Epub 2014 Apr 10. Annu Rev Microbiol. 2014. PMID: 24742054 Free PMC article. Review.
-
Fine structure of the vaccinia virion determined by controlled degradation and immunolocalization.Virology. 2015 Jan 15;475:204-18. doi: 10.1016/j.virol.2014.11.020. Epub 2014 Dec 8. Virology. 2015. PMID: 25486587 Free PMC article.
References
-
- Moss B. Poxviridae: The Viruses and Their Replication. In: Knipe DM, Howley PM, editor. Fields Virology. Lippincott Williams & Wilkins, Phila; 2007. pp. 2905–2946.
-
- Smith GL, Vanderplasschen A. Extracellular enveloped vaccinia virus. Entry, egress, and evasion. Adv Exp Med Biol. 1998;440:395–414. - PubMed
-
- McFadden G, Graham K, Barry M. New strategies of immune modulation by DNA viruses. Transplant Proc. 1996;28:2085–2088. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
