

Reduction of DILP2 in Drosophila triages a metabolic phenotype from lifespan revealing redundancy and compensation among DILPs
- PMID: 19005568
- PMCID: PMC2579582
- DOI: 10.1371/journal.pone.0003721
Reduction of DILP2 in Drosophila triages a metabolic phenotype from lifespan revealing redundancy and compensation among DILPs
Abstract
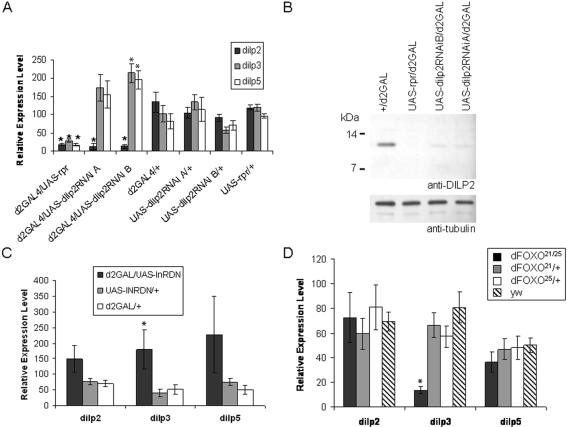
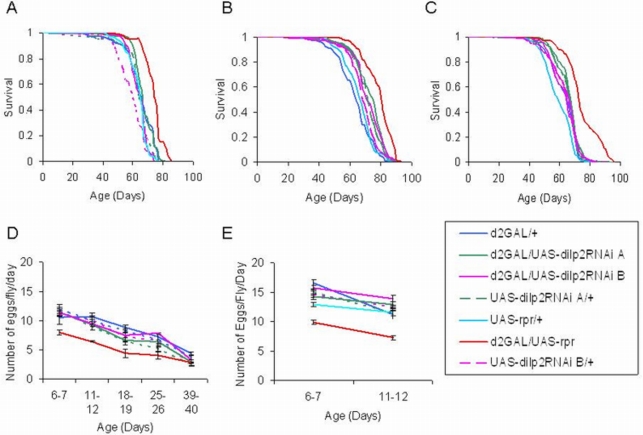
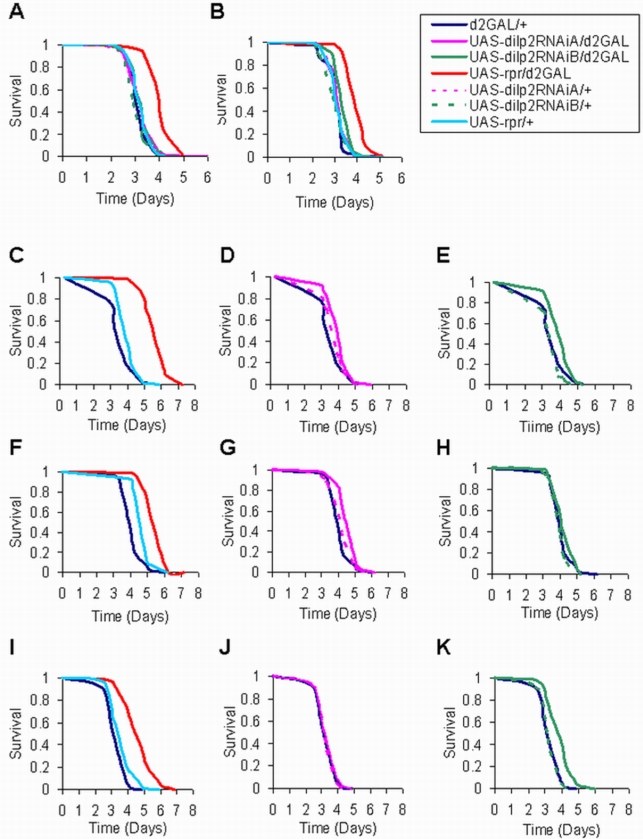
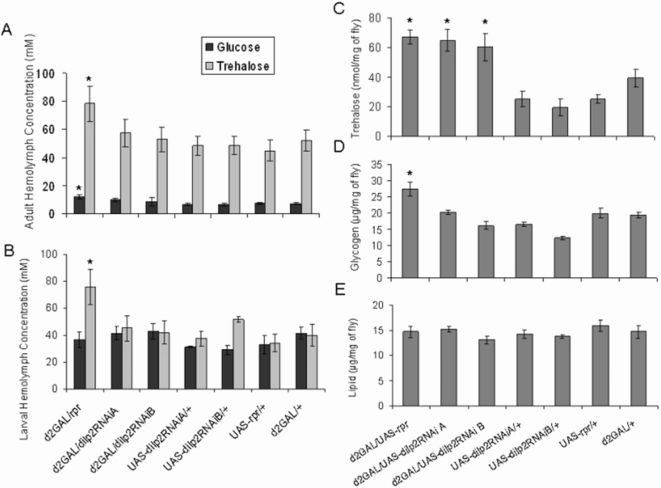
The insulin/IGF-like signalling (IIS) pathway has diverse functions in all multicellular organisms, including determination of lifespan. The seven insulin-like peptides (DILPs) in Drosophila are expressed in a stage- and tissue-specific manner. Partial ablation of the median neurosecretory cells (mNSCs) in the brain, which produce three DILPs, extends lifespan, reduces fecundity, alters lipid and carbohydrate metabolism and increases oxidative stress resistance. To determine if reduced expression of DILPs is causal in these effects, and to investigate possible functional diversification and redundancy between DILPs, we used RNA interference to lower specifically the transcript and protein levels of dilp2, the most highly expressed of the mNSC-derived DILPs. We found that DILP2 was limiting only for the increased whole-body trehalose content associated with mNSC-ablation. We observed a compensatory increase in dilp3 and 5 mRNA upon dilp2 knock down. By manipulation of dfoxo and dInR, we showed that the increase in dilp3 is regulated via autocrine insulin signaling in the mNSCs. Our study demonstrates that, despite the correlation between reduced dilp2 mRNA levels and lifespan-extension often observed, DILP2 reduction is not sufficient to extend lifespan. Nor is the increased trehalose storage associated with reduced IIS sufficient to extend lifespan. To understand the normal regulation of expression of the dilps and any functional diversification between them will require independent control of the expression of different dilps.
Conflict of interest statement
Figures
Similar articles
-
Molecular evolution and functional characterization of Drosophila insulin-like peptides.PLoS Genet. 2010 Feb 26;6(2):e1000857. doi: 10.1371/journal.pgen.1000857. PLoS Genet. 2010. PMID: 20195512 Free PMC article.
-
Lifespan extension by increased expression of the Drosophila homologue of the IGFBP7 tumour suppressor.Aging Cell. 2011 Feb;10(1):137-47. doi: 10.1111/j.1474-9726.2010.00653.x. Aging Cell. 2011. PMID: 21108726 Free PMC article.
-
Factors that regulate expression patterns of insulin-like peptides and their association with physiological and metabolic traits in Drosophila.Insect Biochem Mol Biol. 2021 Aug;135:103609. doi: 10.1016/j.ibmb.2021.103609. Epub 2021 Jun 17. Insect Biochem Mol Biol. 2021. PMID: 34146686 Review.
-
Drosophila insulin-like peptide dilp1 increases lifespan and glucagon-like Akh expression epistatic to dilp2.Aging Cell. 2019 Feb;18(1):e12863. doi: 10.1111/acel.12863. Epub 2018 Dec 3. Aging Cell. 2019. PMID: 30511458 Free PMC article.
-
Functional implications of Drosophila insulin-like peptides in metabolism, aging, and dietary restriction.Front Physiol. 2013 Oct 16;4:288. doi: 10.3389/fphys.2013.00288. Front Physiol. 2013. PMID: 24137131 Free PMC article. Review.
Cited by
-
Complex expression dynamics and robustness in C. elegans insulin networks.Genome Res. 2013 Jun;23(6):954-65. doi: 10.1101/gr.150466.112. Epub 2013 Mar 28. Genome Res. 2013. PMID: 23539137 Free PMC article.
-
Drosophila insulin-like peptide-6 (dilp6) expression from fat body extends lifespan and represses secretion of Drosophila insulin-like peptide-2 from the brain.Aging Cell. 2012 Dec;11(6):978-85. doi: 10.1111/acel.12000. Epub 2012 Sep 18. Aging Cell. 2012. PMID: 22935001 Free PMC article.
-
Molecular evolution and functional characterization of Drosophila insulin-like peptides.PLoS Genet. 2010 Feb 26;6(2):e1000857. doi: 10.1371/journal.pgen.1000857. PLoS Genet. 2010. PMID: 20195512 Free PMC article.
-
A conserved role for syndecan family members in the regulation of whole-body energy metabolism.PLoS One. 2010 Jun 23;5(6):e11286. doi: 10.1371/journal.pone.0011286. PLoS One. 2010. PMID: 20585652 Free PMC article.
-
Positive and negative gustatory inputs affect Drosophila lifespan partly in parallel to dFOXO signaling.Proc Natl Acad Sci U S A. 2014 Jun 3;111(22):8143-8. doi: 10.1073/pnas.1315466111. Epub 2014 May 20. Proc Natl Acad Sci U S A. 2014. PMID: 24847072 Free PMC article.
References
-
- Skorokhod A, Gamulin V, Gundacker D, Kavsan V, Muller IM, et al. Origin of insulin receptor-like tyrosine kinases in marine sponges. Biol Bull. 1999;197:198–206. - PubMed
-
- Butler AA, Le Roith D. Control of growth by the somatropic axis: growth hormone and the insulin-like growth factors have related and independent roles. Annu Rev Physiol. 2001;63:141–164. - PubMed
-
- Brogiolo W, Stocker H, Ikeya T, Rintelen F, Fernandez R, et al. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr Biol. 2001;20;11:213–221. - PubMed
-
- Rulifson EJ, Kim SK, Nusse R. Ablation of insulin-producing neurons in flies: growth and diabetic phenotypes. Science. 2002;296:1118–1120. - PubMed
-
- Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science. 1997;277:942–946. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
