

Meiotic regulation of TPX2 protein levels governs cell cycle progression in mouse oocytes
- PMID: 18833336
- PMCID: PMC2556383
- DOI: 10.1371/journal.pone.0003338
Meiotic regulation of TPX2 protein levels governs cell cycle progression in mouse oocytes
Abstract
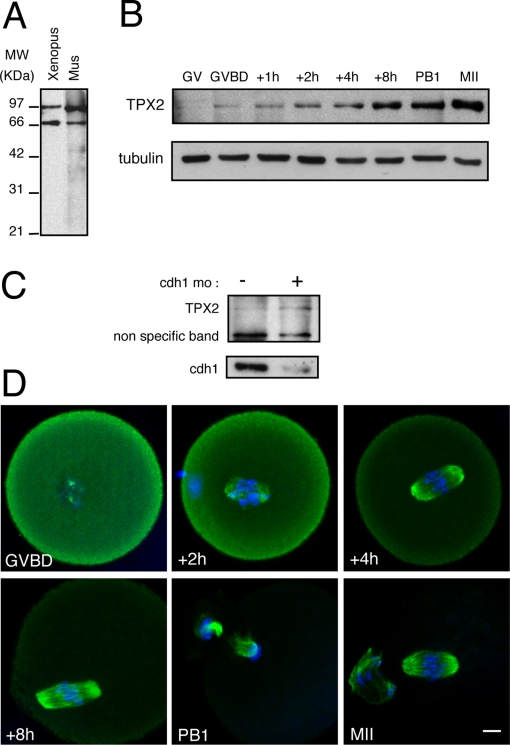
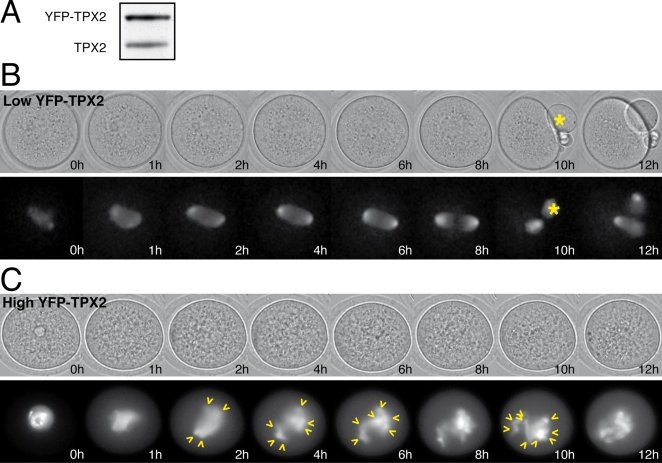
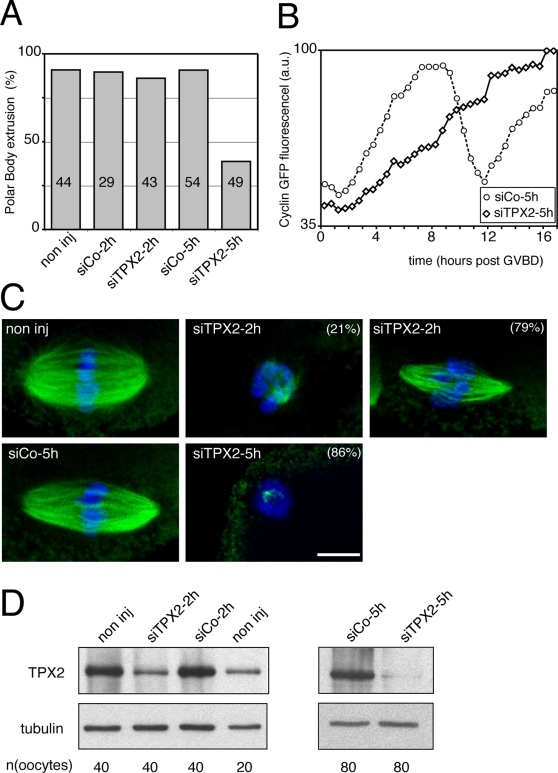
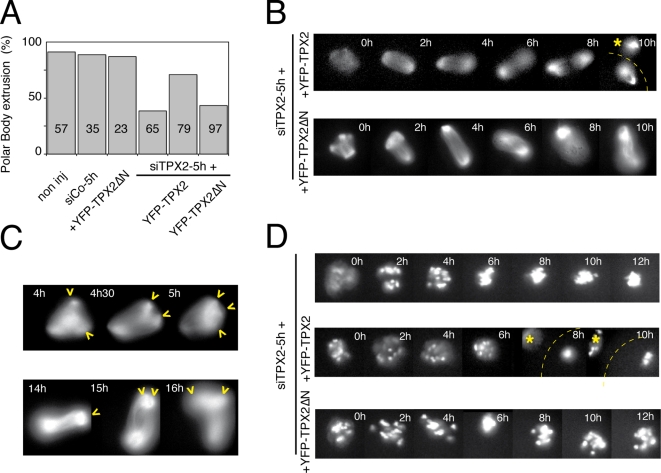
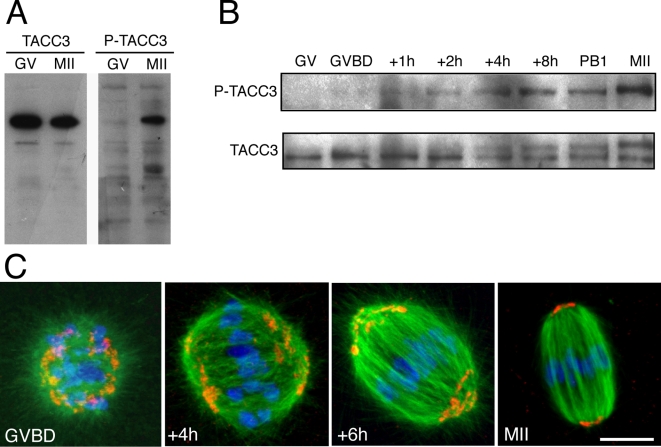
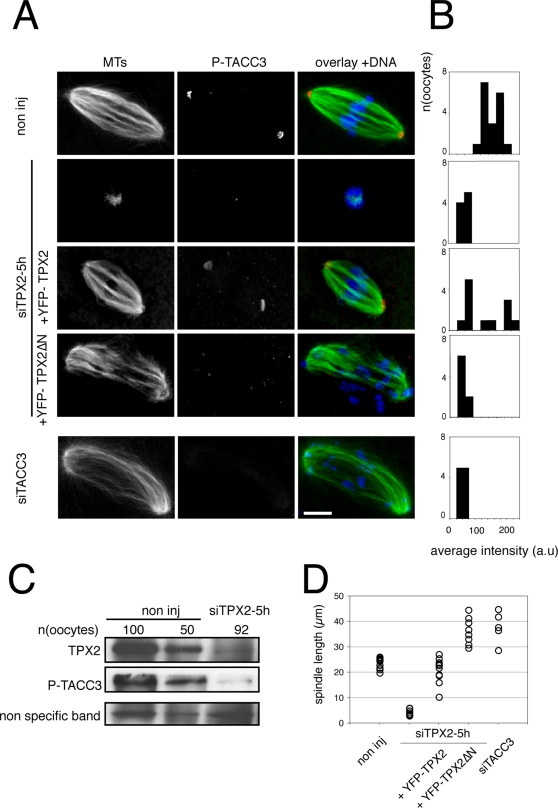
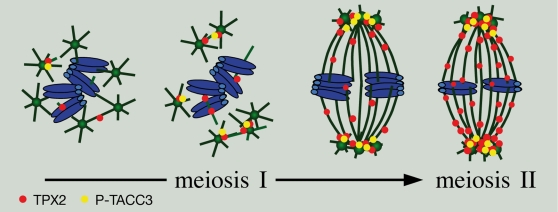
Formation of female gametes requires acentriolar spindle assembly during meiosis. Mitotic spindles organize from centrosomes and via local activation of the RanGTPase on chromosomes. Vertebrate oocytes present a RanGTP gradient centred on chromatin at all stages of meiotic maturation. However, this gradient is dispensable for assembly of the first meiotic spindle. To understand this meiosis I peculiarity, we studied TPX2, a Ran target, in mouse oocytes. Strikingly, TPX2 activity is controlled at the protein level through its accumulation from meiosis I to II. By RNAi depletion and live imaging, we show that TPX2 is required for spindle assembly via two distinct functions. It controls microtubule assembly and spindle pole integrity via the phosphorylation of TACC3, a regulator of MTOCs activity. We show that meiotic spindle formation in vivo depends on the regulation of at least a target of Ran, TPX2, rather than on the regulation of the RanGTP gradient itself.
Conflict of interest statement
Figures

Similar articles
-
A centriole- and RanGTP-independent spindle assembly pathway in meiosis I of vertebrate oocytes.J Cell Biol. 2007 Jan 29;176(3):295-305. doi: 10.1083/jcb.200605199. J Cell Biol. 2007. PMID: 17261848 Free PMC article.
-
Pins homolog LGN regulates meiotic spindle organization in mouse oocytes.Cell Res. 2009 Jul;19(7):838-48. doi: 10.1038/cr.2009.54. Cell Res. 2009. PMID: 19434098
-
Stage specific effects of carbendazim (MBC) on meiotic cell cycle progression in mouse oocytes.Mol Reprod Dev. 1997 Mar;46(3):351-62. doi: 10.1002/(SICI)1098-2795(199703)46:3<351::AID-MRD14>3.0.CO;2-1. Mol Reprod Dev. 1997. PMID: 9041138
-
The role of RanGTP gradient in vertebrate oocyte maturation.Results Probl Cell Differ. 2011;53:235-67. doi: 10.1007/978-3-642-19065-0_12. Results Probl Cell Differ. 2011. PMID: 21630149 Review.
-
Spindle formation, chromosome segregation and the spindle checkpoint in mammalian oocytes and susceptibility to meiotic error.Mutat Res. 2008 Mar 12;651(1-2):14-29. doi: 10.1016/j.mrgentox.2007.10.015. Epub 2007 Nov 9. Mutat Res. 2008. PMID: 18096427 Review.
Cited by
-
Bcl2l10, a new Tpx2 binding partner, is a master regulator of Aurora kinase A in mouse oocytes.Cell Cycle. 2016 Dec;15(23):3296-3305. doi: 10.1080/15384101.2016.1243630. Epub 2016 Oct 18. Cell Cycle. 2016. PMID: 27753540 Free PMC article.
-
Haspin kinase regulates microtubule-organizing center clustering and stability through Aurora kinase C in mouse oocytes.J Cell Sci. 2016 Oct 1;129(19):3648-3660. doi: 10.1242/jcs.189340. Epub 2016 Aug 25. J Cell Sci. 2016. PMID: 27562071 Free PMC article.
-
Curcumin disrupts meiotic and mitotic divisions via spindle impairment and inhibition of CDK1 activity.Cell Prolif. 2010 Aug;43(4):354-64. doi: 10.1111/j.1365-2184.2010.00684.x. Cell Prolif. 2010. PMID: 20590660 Free PMC article.
-
Harnessing nanotopography and integrin-matrix interactions to influence stem cell fate.Nat Mater. 2014 Jun;13(6):558-69. doi: 10.1038/nmat3980. Nat Mater. 2014. PMID: 24845995 Review.
-
Molecular insight into how γ-TuRC makes microtubules.J Cell Sci. 2021 Jul 15;134(14):jcs245464. doi: 10.1242/jcs.245464. Epub 2021 Jul 23. J Cell Sci. 2021. PMID: 34297125 Free PMC article. Review.
References
-
- Eichenlaub-Ritter U. Age-related non-disjunction, spindle formation and progression through maturation of mammalian oocytes. Progress in Clinical and Biological Research. 1989;318:259–269. - PubMed
-
- Hassold T, Hunt P. To err (meiotically) is human: the genesis of human aneuploidy. Nature Review Genetics. 2001;2:280–291. - PubMed
-
- Szöllösi D, Calarco P, Donahue RP. Absence of centrioles in the first and second meiotic spindles of mouse oocytes. Journal of Cell Science. 1972;11:521–541. - PubMed
-
- Schuh M, Ellenberg J. Self-organization of MTOCs replaces centrosome function during acentrosomal spindle assembly in live mouse oocytes. Cell. 2007;130:484–498. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
