

Transforming growth factor-beta suppresses the activation of CD8+ T-cells when naive but promotes their survival and function once antigen experienced: a two-faced impact on autoimmunity
- PMID: 18689691
- PMCID: PMC2551678
- DOI: 10.2337/db08-0609
Transforming growth factor-beta suppresses the activation of CD8+ T-cells when naive but promotes their survival and function once antigen experienced: a two-faced impact on autoimmunity
Abstract
Objective: Transforming growth factor-beta (TGF-beta) can exhibit strong immune suppression but has also been shown to promote T-cell growth. We investigated the differential effect of this cytokine on CD8(+) T-cells in autoimmunity and antiviral immunity.
Research design and methods: We used mouse models for virally induced type 1 diabetes in conjunction with transgenic systems enabling manipulation of TGF-beta expression or signaling in vivo.
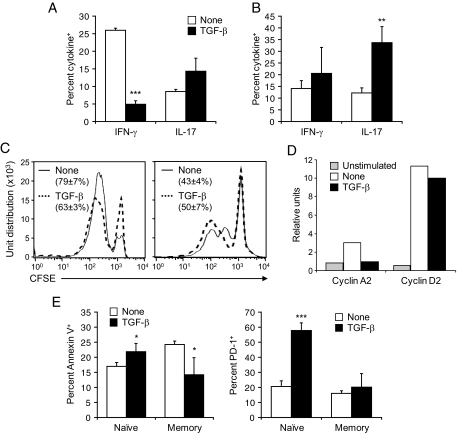
Results: Surprisingly, when expressed selectively in the pancreas, TGF-beta reduced apoptosis of differentiated autoreactive CD8(+) T-cells, favoring their expansion and infiltration of the islets. These results pointed to drastically opposite roles of TGF-beta on naïve compared with antigen-experienced/memory CD8(+) T-cells. Indeed, in the absence of functional TGF-beta signaling in T-cells, fast-onset type 1 diabetes caused by activation of naïve CD8(+) T-cells occurred faster, whereas slow-onset disease depending on accumulation and activation of antigen-experienced/memory CD8(+) T-cells was decreased. TGF-beta receptor-deficient CD8(+) T-cells showed enhanced activation and expansion after lymphocytic choriomeningitis virus infection in vivo but were more prone to apoptosis once antigen experienced and failed to survive as functional memory cells. In vitro, TGF-beta suppressed naïve CD8(+) T-cell activation and gamma-interferon production, whereas memory CD8(+) T-cells stimulated in the presence of TGF-beta showed enhanced survival and increased production of interleukin-17 in conjunction with gamma-interferon.
Conclusions: The effect of TGF-beta on CD8(+) T-cells is dependent on their differentiation status and activation history. These results highlight a novel aspect of the pleiotropic nature of TGF-beta and have implications for the design of immune therapies involving this cytokine.
Figures
Similar articles
-
Clonal restriction of the expansion of antigen-specific CD8+ memory T cells by transforming growth factor-{beta}.J Leukoc Biol. 2006 May;79(5):1033-42. doi: 10.1189/jlb.0805474. Epub 2006 Feb 14. J Leukoc Biol. 2006. PMID: 16478921
-
Infiltration of tumor-reactive transforming growth factor-beta insensitive CD8+ T cells into the tumor parenchyma is associated with apoptosis and rejection of tumor cells.Prostate. 2006 Feb 15;66(3):235-47. doi: 10.1002/pros.20340. Prostate. 2006. PMID: 16173028
-
Krüppel-like factor KLF10 regulates transforming growth factor receptor II expression and TGF-β signaling in CD8+ T lymphocytes.Am J Physiol Cell Physiol. 2015 Mar 1;308(5):C362-71. doi: 10.1152/ajpcell.00262.2014. Epub 2014 Dec 3. Am J Physiol Cell Physiol. 2015. PMID: 25472963 Free PMC article.
-
Efficacy Against Human Prostate Cancer by Prostate-specific Membrane Antigen-specific, Transforming Growth Factor-β Insensitive Genetically Targeted CD8+ T-cells Derived from Patients with Metastatic Castrate-resistant Disease.Eur Urol. 2018 May;73(5):648-652. doi: 10.1016/j.eururo.2017.12.008. Epub 2017 Dec 21. Eur Urol. 2018. PMID: 29275833 Free PMC article. Review.
-
TGF-β Regulation of T Cells.Annu Rev Immunol. 2023 Apr 26;41:483-512. doi: 10.1146/annurev-immunol-101921-045939. Epub 2023 Feb 7. Annu Rev Immunol. 2023. PMID: 36750317 Review.
Cited by
-
TGF-β and IL-23 gene expression in unstimulated PBMCs of patients with diabetes.Endocrine. 2012 Jun;41(3):430-4. doi: 10.1007/s12020-011-9578-7. Epub 2011 Dec 17. Endocrine. 2012. PMID: 22180056
-
TGF-β in T Cell Biology: Implications for Cancer Immunotherapy.Cancers (Basel). 2018 Jun 11;10(6):194. doi: 10.3390/cancers10060194. Cancers (Basel). 2018. PMID: 29891791 Free PMC article. Review.
-
TGF-beta enhances effector Th1 cell activation but promotes self-regulation via IL-10.J Immunol. 2010 May 15;184(10):5628-36. doi: 10.4049/jimmunol.1000288. Epub 2010 Apr 14. J Immunol. 2010. PMID: 20393141 Free PMC article.
-
TGF-β, Bone Morphogenetic Protein, and Activin Signaling and the Tumor Microenvironment.Cold Spring Harb Perspect Biol. 2017 May 1;9(5):a022285. doi: 10.1101/cshperspect.a022285. Cold Spring Harb Perspect Biol. 2017. PMID: 28062564 Free PMC article. Review.
-
Dysregulation of serum gamma interferon levels in vascular chronic Q Fever patients provides insights into disease pathogenesis.Clin Vaccine Immunol. 2015 Jun;22(6):664-71. doi: 10.1128/CVI.00078-15. Epub 2015 Apr 29. Clin Vaccine Immunol. 2015. PMID: 25924761 Free PMC article.
References
-
- Gorelik L, Flavell RA: Transforming growth factor-beta in T-cell biology. Nat Rev Immunol 2:46–53, 2002 - PubMed
-
- Gorelik L, Flavell RA: Abrogation of TGFbeta signaling in T cells leads to spontaneous T cell differentiation and autoimmune disease. Immunity 12:171–181, 2000 - PubMed
-
- Letterio JJ, Roberts AB: Regulation of immune responses by TGF-beta. Annu Rev Immunol 16:137–161, 1998 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
