

Arabidopsis protein disulfide isomerase-5 inhibits cysteine proteases during trafficking to vacuoles before programmed cell death of the endothelium in developing seeds
- PMID: 18676877
- PMCID: PMC2553623
- DOI: 10.1105/tpc.108.058339
Arabidopsis protein disulfide isomerase-5 inhibits cysteine proteases during trafficking to vacuoles before programmed cell death of the endothelium in developing seeds
Abstract
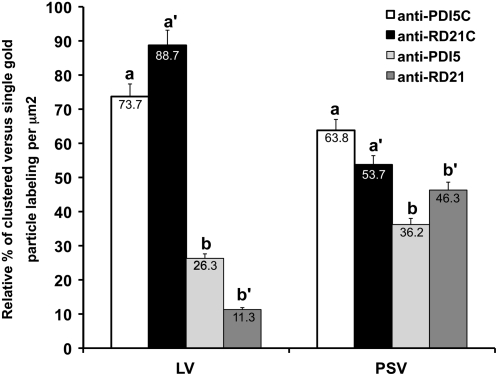
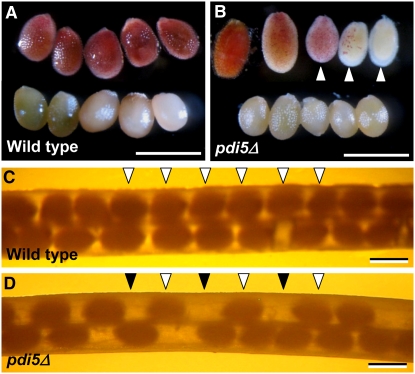
Protein disulfide isomerase (PDI) oxidizes, reduces, and isomerizes disulfide bonds, modulates redox responses, and chaperones proteins. The Arabidopsis thaliana genome contains 12 PDI genes, but little is known about their subcellular locations and functions. We demonstrate that PDI5 is expressed in endothelial cells about to undergo programmed cell death (PCD) in developing seeds. PDI5 interacts with three different Cys proteases in yeast two-hybrid screens. One of these traffics together with PDI5 from the endoplasmic reticulum through the Golgi to vacuoles, and its recombinant form is functionally inhibited by recombinant PDI5 in vitro. Peak PDI5 expression in endothelial cells precedes PCD, whereas decreasing PDI5 levels coincide with the onset of PCD-related cellular changes, such as enlargement and subsequent collapse of protein storage vacuoles, lytic vacuole shrinkage and degradation, and nuclear condensation and fragmentation. Loss of PDI5 function leads to premature initiation of PCD during embryogenesis and to fewer, often nonviable, seeds. We propose that PDI5 is required for proper seed development and regulates the timing of PCD by chaperoning and inhibiting Cys proteases during their trafficking to vacuoles before PCD of the endothelial cells. During this transitional phase of endothelial cell development, the protein storage vacuoles become the de facto lytic vacuoles that mediate PCD.
Figures
Similar articles
-
Protein disulfide isomerase-2 of Arabidopsis mediates protein folding and localizes to both the secretory pathway and nucleus, where it interacts with maternal effect embryo arrest factor.Mol Cells. 2011 Nov;32(5):459-75. doi: 10.1007/s10059-011-0150-3. Epub 2011 Sep 5. Mol Cells. 2011. PMID: 21909944 Free PMC article.
-
At14a-Like1 participates in membrane-associated mechanisms promoting growth during drought in Arabidopsis thaliana.Proc Natl Acad Sci U S A. 2015 Aug 18;112(33):10545-50. doi: 10.1073/pnas.1510140112. Epub 2015 Aug 3. Proc Natl Acad Sci U S A. 2015. PMID: 26240315 Free PMC article.
-
An ER-localized form of PV72, a seed-specific vacuolar sorting receptor, interferes the transport of an NPIR-containing proteinase in Arabidopsis leaves.Plant Cell Physiol. 2004 Jan;45(1):9-17. doi: 10.1093/pcp/pch012. Plant Cell Physiol. 2004. PMID: 14749481
-
Programmed cell death in Ricinus and Arabidopsis: the function of KDEL cysteine peptidases in development.Physiol Plant. 2012 May;145(1):103-13. doi: 10.1111/j.1399-3054.2012.01580.x. Epub 2012 Feb 17. Physiol Plant. 2012. PMID: 22268582 Review.
-
Endomembrane-mediated storage protein trafficking in plants: Golgi-dependent or Golgi-independent?FEBS Lett. 2022 Sep;596(17):2215-2230. doi: 10.1002/1873-3468.14374. Epub 2022 Jun 10. FEBS Lett. 2022. PMID: 35615915 Review.
Cited by
-
Flower bud proteome reveals modulation of sex-biased proteins potentially associated with sex expression and modification in dioecious Coccinia grandis.BMC Plant Biol. 2019 Jul 23;19(1):330. doi: 10.1186/s12870-019-1937-1. BMC Plant Biol. 2019. PMID: 31337343 Free PMC article.
-
Pull the fuzes: Processing protein precursors to generate apoplastic danger signals for triggering plant immunity.Plant Commun. 2024 Aug 12;5(8):100931. doi: 10.1016/j.xplc.2024.100931. Epub 2024 Apr 30. Plant Commun. 2024. PMID: 38689495 Free PMC article. Review.
-
P5-type sulfhydryl oxidoreductase promotes the sorting of proteins to protein body I in rice endosperm cells.Plant Signal Behav. 2013 Feb;8(2):e23075. doi: 10.4161/psb.23075. Epub 2013 Jan 8. Plant Signal Behav. 2013. PMID: 23299424 Free PMC article.
-
The Arabidopsis RING E3 ubiquitin ligase AtAIRP3/LOG2 participates in positive regulation of high-salt and drought stress responses.Plant Physiol. 2013 Jul;162(3):1733-49. doi: 10.1104/pp.113.220103. Epub 2013 May 21. Plant Physiol. 2013. PMID: 23696092 Free PMC article.
-
New Arabidopsis thaliana cytochrome c partners: a look into the elusive role of cytochrome c in programmed cell death in plants.Mol Cell Proteomics. 2013 Dec;12(12):3666-76. doi: 10.1074/mcp.M113.030692. Epub 2013 Sep 9. Mol Cell Proteomics. 2013. PMID: 24019145 Free PMC article.
References
-
- Andème-Onzighi, C., Sivaguru, M., Judy-March, J., Baskin, T.I., and Driouich, A. (2002). The reb1-1 mutation of Arabidopsis alters the morphology of trichoblasts, the expression of arabinogalactan-proteins and the organization of cortical microtubules. Planta 215 949–958. - PubMed
-
- Beeckman, T., Rycke, R.D., Viane, R., and Inzé, D. (2000). Histological study of seed coat development in Arabidopsis thaliana. J. Plant Res. 113 139–148.
-
- Beers, E.P., Woffenden, B.J., and Zhao, C. (2000). Plant proteolytic enzymes: Possible roles during programmed cell death. Plant Mol. Biol. 44 399–415. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
