

An exocyst complex functions in plant cell growth in Arabidopsis and tobacco
- PMID: 18492870
- PMCID: PMC2438459
- DOI: 10.1105/tpc.108.059105
An exocyst complex functions in plant cell growth in Arabidopsis and tobacco
Abstract
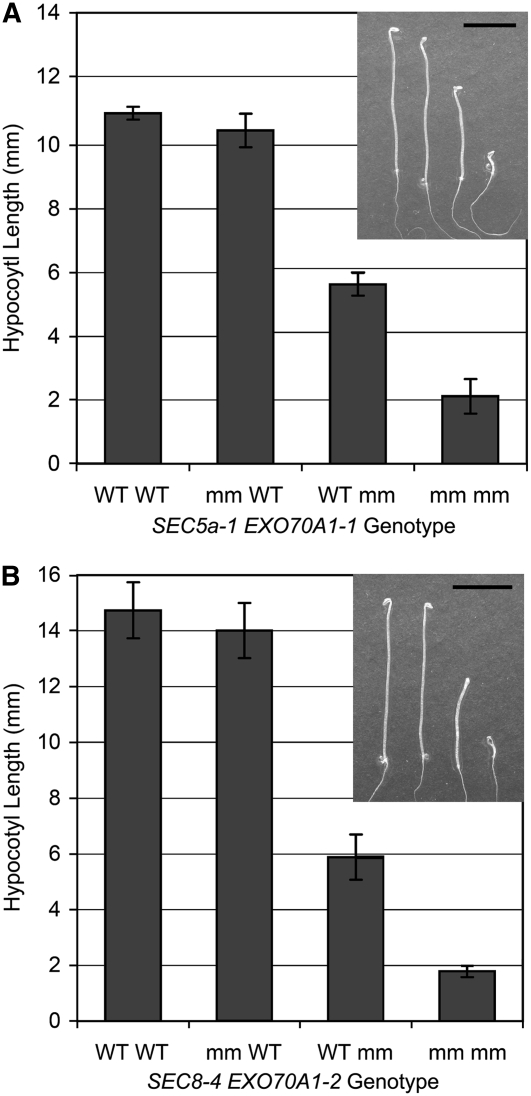
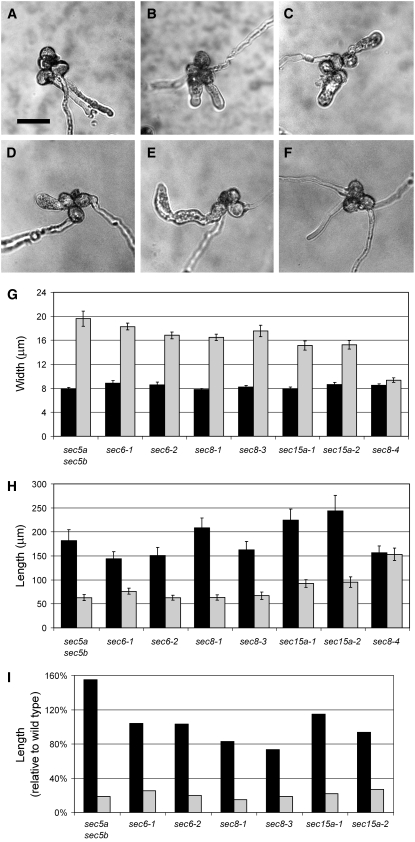
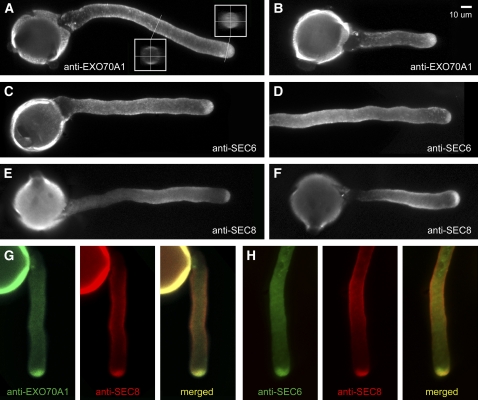
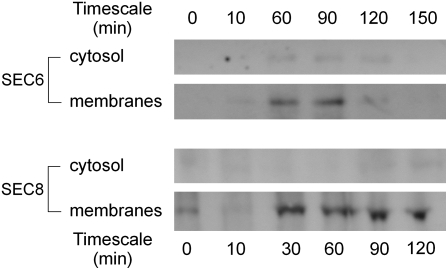
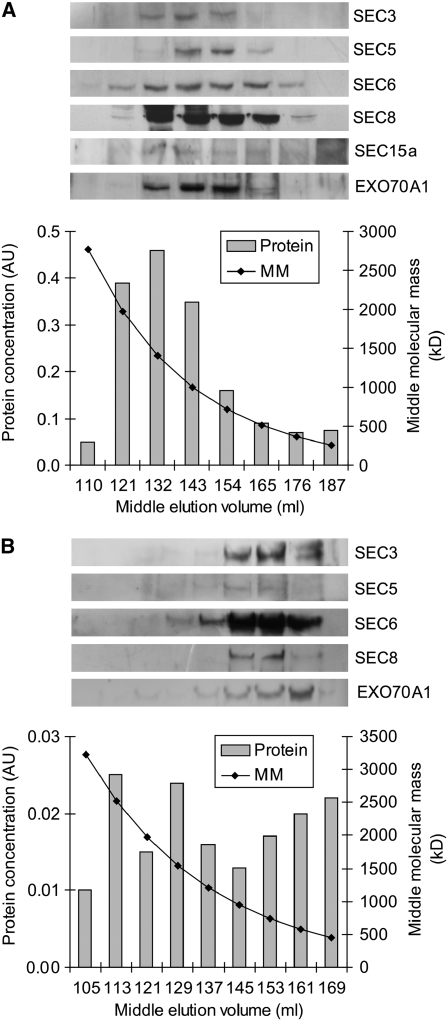
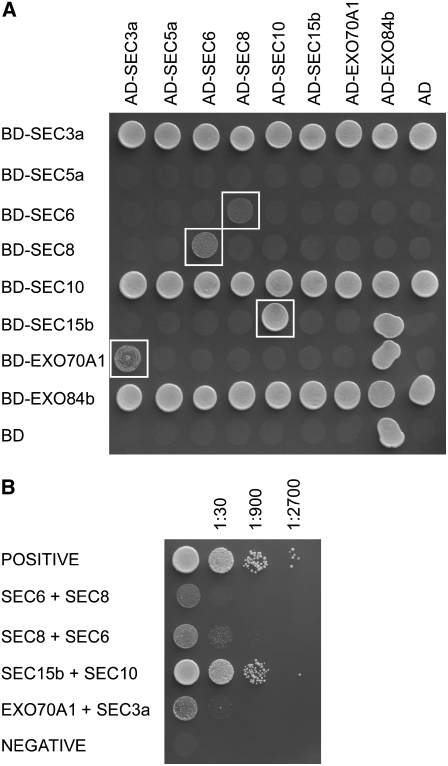
The exocyst, an octameric tethering complex and effector of Rho and Rab GTPases, facilitates polarized secretion in yeast and animals. Recent evidence implicates three plant homologs of exocyst subunits (SEC3, SEC8, and EXO70A1) in plant cell morphogenesis. Here, we provide genetic, cell biological, and biochemical evidence that these and other predicted subunits function together in vivo in Arabidopsis thaliana. Double mutants in exocyst subunits (sec5 exo70A1 and sec8 exo70A1) show a synergistic defect in etiolated hypocotyl elongation. Mutants in exocyst subunits SEC5, SEC6, SEC8, and SEC15a show defective pollen germination and pollen tube growth phenotypes. Using antibodies directed against SEC6, SEC8, and EXO70A1, we demonstrate colocalization of these proteins at the apex of growing tobacco pollen tubes. The SEC3, SEC5, SEC6, SEC8, SEC10, SEC15a, and EXO70 subunits copurify in a high molecular mass fraction of 900 kD after chromatographic fractionation of an Arabidopsis cell suspension extract. Blue native electrophoresis confirmed the presence of SEC3, SEC6, SEC8, and EXO70 in high molecular mass complexes. Finally, use of the yeast two-hybrid system revealed interaction of Arabidopsis SEC3a with EXO70A1, SEC10 with SEC15b, and SEC6 with SEC8. We conclude that the exocyst functions as a complex in plant cells, where it plays important roles in morphogenesis.
Figures
Comment in
-
An exocyst vesicle tethering complex in plants.Plant Cell. 2008 May;20(5):1188. doi: 10.1105/tpc.108.200511. Epub 2008 May 20. Plant Cell. 2008. PMID: 18492869 Free PMC article. No abstract available.
Similar articles
-
Exocyst SEC3 and Phosphoinositides Define Sites of Exocytosis in Pollen Tube Initiation and Growth.Plant Physiol. 2016 Oct;172(2):980-1002. doi: 10.1104/pp.16.00690. Epub 2016 Aug 11. Plant Physiol. 2016. PMID: 27516531 Free PMC article.
-
An exocyst vesicle tethering complex in plants.Plant Cell. 2008 May;20(5):1188. doi: 10.1105/tpc.108.200511. Epub 2008 May 20. Plant Cell. 2008. PMID: 18492869 Free PMC article. No abstract available.
-
EXO70C2 Is a Key Regulatory Factor for Optimal Tip Growth of Pollen.Plant Physiol. 2017 May;174(1):223-240. doi: 10.1104/pp.16.01282. Epub 2017 Mar 29. Plant Physiol. 2017. PMID: 28356503 Free PMC article.
-
Diverse Functions and Signal Transduction of the Exocyst Complex in Tumor Cells.J Cell Physiol. 2017 May;232(5):939-957. doi: 10.1002/jcp.25619. Epub 2016 Dec 22. J Cell Physiol. 2017. PMID: 27669116 Review.
-
The exocyst complex in polarized exocytosis.Int Rev Cytol. 2004;233:243-65. doi: 10.1016/S0074-7696(04)33006-8. Int Rev Cytol. 2004. PMID: 15037366 Review.
Cited by
-
Arabidopsis COG Complex Subunits COG3 and COG8 Modulate Golgi Morphology, Vesicle Trafficking Homeostasis and Are Essential for Pollen Tube Growth.PLoS Genet. 2016 Jul 22;12(7):e1006140. doi: 10.1371/journal.pgen.1006140. eCollection 2016 Jul. PLoS Genet. 2016. PMID: 27448097 Free PMC article.
-
Signalling Pinpointed to the Tip: The Complex Regulatory Network That Allows Pollen Tube Growth.Plants (Basel). 2020 Aug 26;9(9):1098. doi: 10.3390/plants9091098. Plants (Basel). 2020. PMID: 32859043 Free PMC article. Review.
-
Exorcising the exocyst complex.Traffic. 2012 Jul;13(7):898-907. doi: 10.1111/j.1600-0854.2012.01353.x. Epub 2012 Apr 8. Traffic. 2012. PMID: 22420621 Free PMC article. Review.
-
Tethering of Multi-Vesicular Bodies and the Tonoplast to the Plasma Membrane in Plants.Front Plant Sci. 2019 May 22;10:636. doi: 10.3389/fpls.2019.00636. eCollection 2019. Front Plant Sci. 2019. PMID: 31396242 Free PMC article.
-
Expression and functional analyses of EXO70 genes in Arabidopsis implicate their roles in regulating cell type-specific exocytosis.Plant Physiol. 2010 Dec;154(4):1819-30. doi: 10.1104/pp.110.164178. Epub 2010 Oct 13. Plant Physiol. 2010. PMID: 20943851 Free PMC article.
References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657. - PubMed
-
- Andrews, H.K., Zhang, Y.Q., Trotta, N., and Broadie, K. (2002). Drosophila Sec10 is required for hormone secretion but not general exocytosis or neurotransmission. Traffic 12 906–921. - PubMed
-
- Berghöfer, J., and Klösgen, R.B. (1999). Two distinct translocation intermediates can be distinguished during protein transport by the TAT (Deltaph) pathway across the thylakoid membrane. FEBS Lett. 460 328–332. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
