

Review
doi: 10.1038/nri2318.
Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections
Affiliations
- PMID: 18469829
- PMCID: PMC5573867
- DOI: 10.1038/nri2318
Item in Clipboard
Review
Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections
Nat Rev Immunol.
2008 Jul.
Abstract
A diverse and well-balanced repertoire of T cells is thought to be crucial for the efficacious defence against infection with new or re-emerging pathogens throughout life. In the last third of the mammalian lifespan, the maintenance of a balanced T-cell repertoire becomes highly challenging because of the changes in T-cell production and consumption. In this Review, I question whether latent persistent pathogens might be key factors that drive this imbalance and whether they determine the extent of age-associated immune deficiency.
Figures
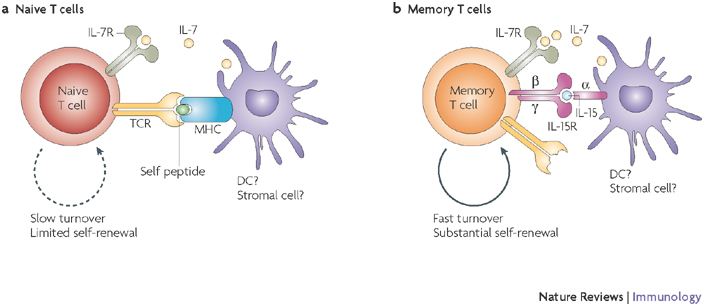
Key factors important for survival and maintenance of naive (a) and memory (b) T cells are summarized. Interleukin-7 (IL-7; for naive T cells and to a lesser extent for memory T cells), IL-15 (for memory T cells) and self-peptide–MHC complexes (for naive T cells) are known to be the key factors for the maintenance of different T-cell subsets. The cellular source of these signals is not defined, but could regulate the overall T-cell numbers by restricted physical distribution and/or numbers. The rate and the extent of self-renewal are different for naive (low) and memory (high) T cells. Question marks indicate speculative or insufficiently documented cellular or molecular interactions. R, receptor; TCR, T-cell receptor.
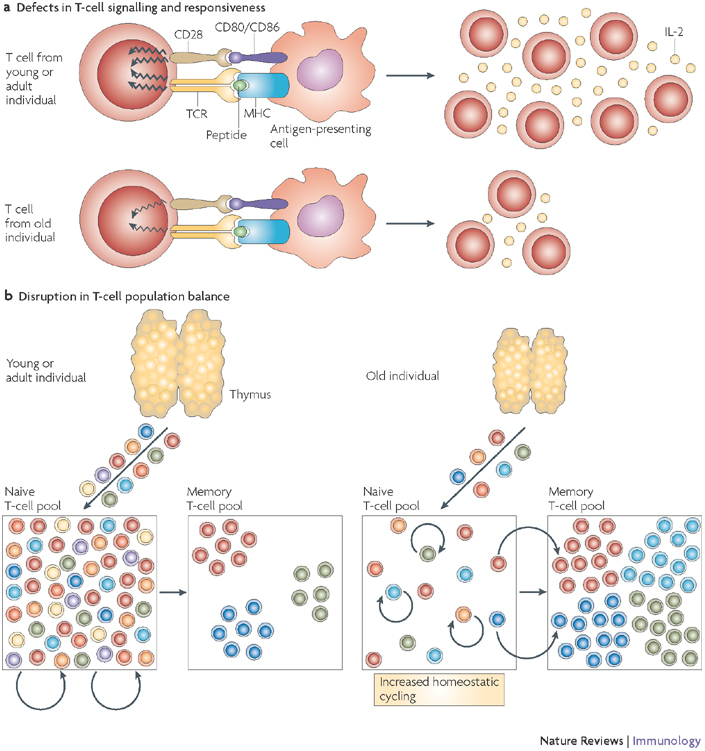
a | A decline in the T-cell response to stimulation through the T-cell receptor (TCR) and co-stimulatory pathways occurs frequently with ageing and is typically shown in ex vivo experiments using agonist T-cell ligands in the absence of antigen-presenting cells, thus showing a cell-autonomous T-cell defect. These proximal signalling defects are followed by a marked decrease in interleukin-2 (IL-2) secretion and T-cell proliferation. b | T-cell population changes result from decreased T-cell production from an involuted thymus, which critically contributes to the depletion of the naive T-cell pool. The naive T-cell pool then undergoes increased homeostatic cycling. This maintains the naive compartment for a while, but as the increased cycling (approximately once every month in mice) turns into homeostatic proliferation (four or more divisions per month, at or above the level seen in the T-cell memory pool),, phenotypic conversion of naive T cells into memory phenotype cells has the potential to further deplete the naive T-cell compartment. This, and the conditions favourable to disrupt maintenance of the memory compartment, provide impetus for T-cell clonal expansions.
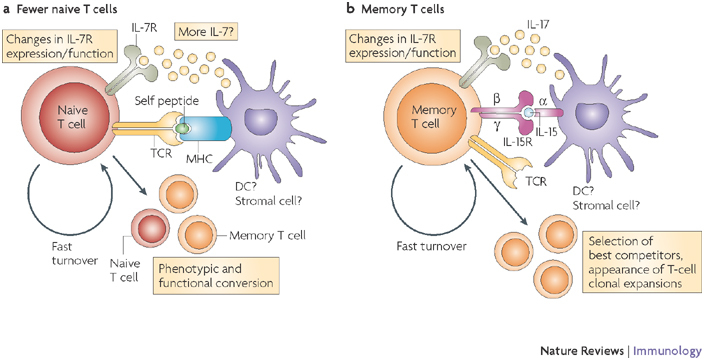
Although the thymus in young individuals produces sufficient numbers of naive T cells that express diverse T-cell receptors (TCRs) and that only infrequently cycle, the involuting, ageing thymus produces many fewer naive T cells (a) and these cells survive poorly in aged individuals. It would be expected that fewer naive T cells would result in less consumption of interleukin-7 (IL-7) and therefore that more IL-7 would be available. This is on the proviso that IL-7 production is not affected with ageing, which remains speculative. Other known age-related alterations include altered expression and function of the IL-7 receptor (IL-7R), and increased turnover of naive T cells,, which has the potential to precipitate their conversion into memory T cells (b). This process would also support the formation of antigen-independent T-cell clonal expansions,. Memory T cells are known to proportionally and numerically increase with ageing,,, and several processes contribute to this increase: loss of naive T cells and increased availability of and/or responsiveness to T-cell maintenance cytokines, such as IL-7 and IL-15. Other factors (not shown), including an increase in the levels of pro-inflammatory cytokines (such as IL-6 and tumour-necrosis factor) and changes in systemic and local levels of hormones (insulin-like growth factor 1, growth hormone, ghrelin and leptin) could also have a role, although this is yet to be proven. DC, dendritic cell.

Stages of the immune response to viruses that establish acute (a) and persistent (b) infection. In both cases, the kinetics of activation and waning of innate immunity, the kinetics of infectious virus and the kinetics of the initial, primary T-cell response are essentially the same. Key differences between acute and persistent viral infections emerge subsequently: the causative agent of acute infection is eliminated and T-cell memory is therefore maintained at a more or less constant, low level, whereas latent persistent pathogens will periodically reactivate, each time then activating the innate immune system. Because innate immunity does not have the property of memory, innate cells do not accumulate. However, memory T cells accumulate following each stimulation and therefore produce memory inflation. This process is pronounced for CD8+ T cells and is more discrete for CD4+ T cells.

Relative availability of, and therefore the exposure of T cells to, viral antigens in the course of infection with different persistent viruses. Latent viruses reach fairly high antigen levels in the course of primary infection but then establish latency and reactivate intermittently, with different frequencies (cytomegalovirus (CMV) more often than herpes simplex virus (HSV)). By contrast, HIV antigens, representative of a chronic persistent virus infection, are present at very high levels initially, but importantly, they also remain at high levels in the blood during the set-point of infection. This provides T cells with constant stimulation, and eventually leads to exhaustion. Note that this illustration is mostly based on indirect immunological parameters for CMV and HSV, rather than on direct identification of the virus or viral antigen, which is difficult to achieve during the persistent phase of the infection.
Similar articles
-
Modelling naive T-cell homeostasis: consequences of heritable cellular lifespan during ageing.Immunol Cell Biol. 2009 Aug-Sep;87(6):445-56. doi: 10.1038/icb.2009.11. Epub 2009 Mar 17. Immunol Cell Biol. 2009. PMID: 19290017
-
The golden anniversary of the thymus.Nat Rev Immunol. 2011 May 27;11(7):489-95. doi: 10.1038/nri2993. Nat Rev Immunol. 2011. PMID: 21617694 Review.
-
T cell homeostasis in centenarians: from the thymus to the periphery.Curr Pharm Des. 2010;16(6):597-603. doi: 10.2174/138161210790883705. Curr Pharm Des. 2010. PMID: 20388069 Review.
-
Comment on "Cutting edge: human CD4-CD8- thymocytes express FOXP3 in the absence of a TCR".J Immunol. 2008 Jul 15;181(2):857-8; author reply 858. doi: 10.4049/jimmunol.181.2.857. J Immunol. 2008. PMID: 18606635 No abstract available.
-
Alterations in the Thymic Selection Threshold Skew the Self-Reactivity of the TCR Repertoire in Neonates.J Immunol. 2017 Aug 1;199(3):965-973. doi: 10.4049/jimmunol.1602137. Epub 2017 Jun 28. J Immunol. 2017. PMID: 28659353
Cited by
-
Chemotherapy combined with immune checkpoint inhibitors may overcome the detrimental effect of high neutrophil-to-lymphocyte ratio prior to treatment in esophageal cancer patients.Front Oncol. 2024 Oct 11;14:1449941. doi: 10.3389/fonc.2024.1449941. eCollection 2024. Front Oncol. 2024. PMID: 39464714 Free PMC article.
-
Age-related epithelial defects limit thymic function and regeneration.Nat Immunol. 2024 Sep;25(9):1593-1606. doi: 10.1038/s41590-024-01915-9. Epub 2024 Aug 7. Nat Immunol. 2024. PMID: 39112630 Free PMC article.
-
In vivo efficacy of phage cocktails against carbapenem resistance Acinetobacter baumannii in the rat pneumonia model.J Virol. 2024 Jul 23;98(7):e0046724. doi: 10.1128/jvi.00467-24. Epub 2024 Jun 12. J Virol. 2024. PMID: 38864621
-
The aged tumor microenvironment limits T cell control of cancer.Nat Immunol. 2024 Jun;25(6):1033-1045. doi: 10.1038/s41590-024-01828-7. Epub 2024 May 14. Nat Immunol. 2024. PMID: 38745085 Free PMC article.
-
Anti-cytomegalovirus antibody levels stratify human immune profiles across the lifespan.Geroscience. 2024 Oct;46(5):4225-4242. doi: 10.1007/s11357-024-01124-0. Epub 2024 Mar 21. Geroscience. 2024. PMID: 38512581 Free PMC article.
References
-
- Brody JA, Brock DB. Handbook of the Biology of Aging. 1985. Epidemiologic and statistical characteristics of the United States elderly population; pp. 3–42.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
