

Role of the translational repressor 4E-BP1 in the regulation of p21(Waf1/Cip1) expression by retinoids
- PMID: 18280804
- PMCID: PMC2390818
- DOI: 10.1016/j.bbrc.2008.02.028
Role of the translational repressor 4E-BP1 in the regulation of p21(Waf1/Cip1) expression by retinoids
Abstract
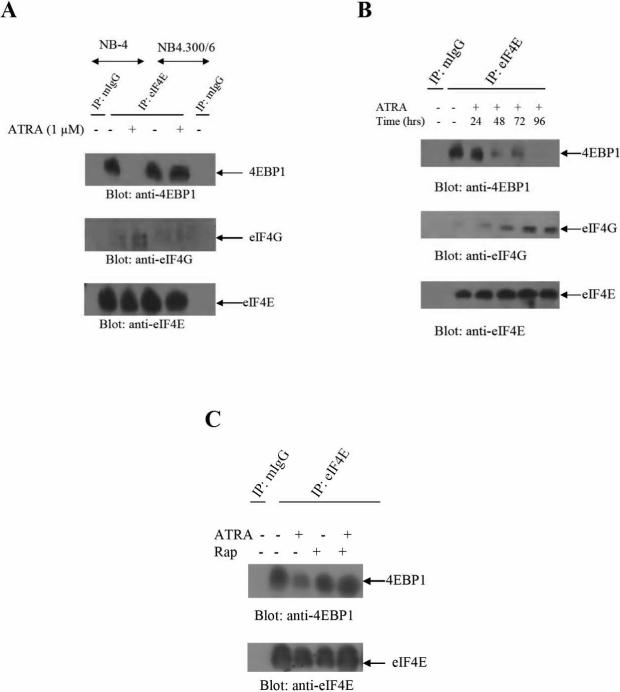
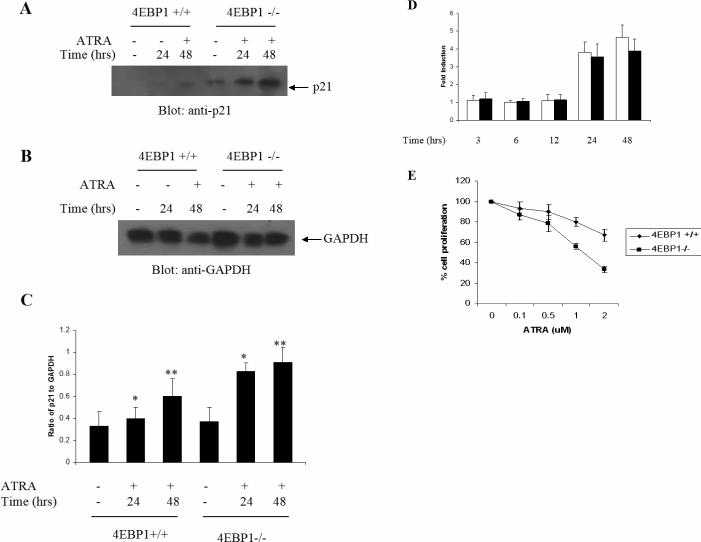
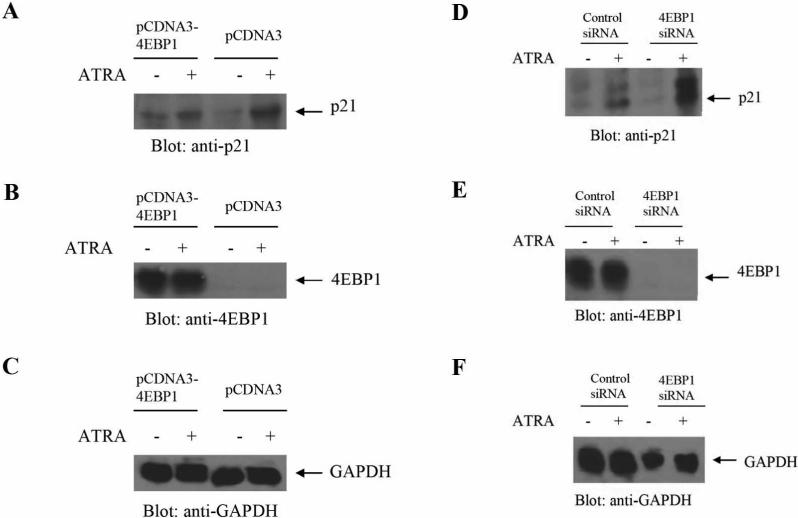
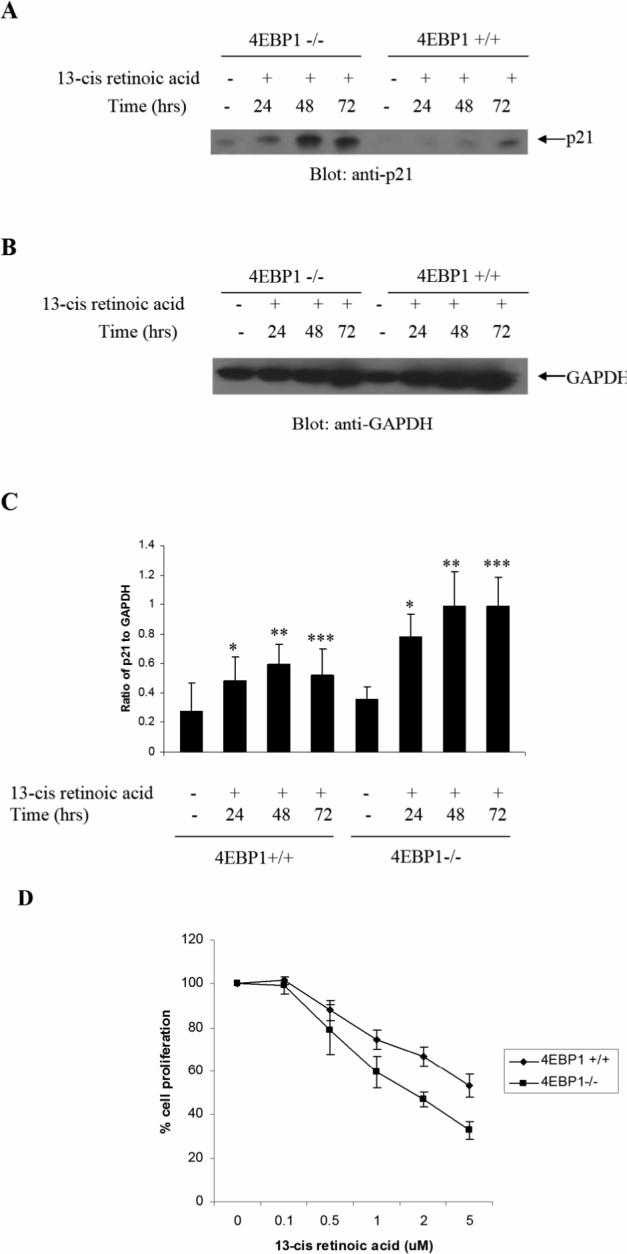
The mechanisms by which retinoids regulate initiation of mRNA translation for proteins that mediate their biological effects are not known. We have previously shown that all-trans-retinoic acid (ATRA) induces mTOR-mediated activation of the p70 S6 kinase, suggesting the existence of a mechanism by which retinoids may regulate mRNA translation. We now demonstrate that treatment of acute promyelocytic leukemia (APL)-derived NB4 cells with ATRA results in dissociation of the translational repressor 4E-BP1 from the eukaryotic initiation factor eIF4E, and subsequent formation of eIF4G-eIF4E complexes. We also show that siRNA-mediated inhibition of 4E-BP1 expression enhances ATRA-dependent upregulation of p21(Waf1/Cip1), a protein that plays a key role in the induction of retinoid-dependent responses. Our data also establish that ATRA- or cis-RA-dependent p21(Waf1/Cip1) protein expression is enhanced in mouse embryonic fibroblasts with targeted disruption of the 4e-bp1 gene, in the absence of any effects on the transcriptional regulation of the p21(Waf1/Cip1) gene. Moreover, generation of ATRA- or cis-retinoic acid (cis-RA)-antiproliferative responses is enhanced in 4E-BP1 knockout cells. Altogether, these findings strongly suggest a key regulatory role for the translational repressor 4E-BP1 in the generation of retinoid-dependent functional responses.
Figures
Similar articles
-
Defect in the regulation of 4E-BP1 and 2, two repressors of translation initiation, in the retinoid acid resistant cell lines, NB4-R1 and NB4-R2.Leukemia. 2000 Nov;14(11):1909-14. doi: 10.1038/sj.leu.2401904. Leukemia. 2000. PMID: 11069026
-
1,25-Dihydroxyvitamin D3 induces differentiation of a retinoic acid-resistant acute promyelocytic leukemia cell line (UF-1) associated with expression of p21(WAF1/CIP1) and p27(KIP1).Blood. 1999 Apr 1;93(7):2225-33. Blood. 1999. PMID: 10090931
-
Activation of the p70 S6 kinase by all-trans-retinoic acid in acute promyelocytic leukemia cells.Blood. 2005 Feb 15;105(4):1669-77. doi: 10.1182/blood-2004-06-2078. Epub 2004 Oct 7. Blood. 2005. PMID: 15471950
-
4E-BP1, a multifactor regulated multifunctional protein.Cell Cycle. 2016;15(6):781-6. doi: 10.1080/15384101.2016.1151581. Cell Cycle. 2016. PMID: 26901143 Free PMC article. Review.
-
The Role of LncRNAs in Translation.Noncoding RNA. 2021 Feb 20;7(1):16. doi: 10.3390/ncrna7010016. Noncoding RNA. 2021. PMID: 33672592 Free PMC article. Review.
Cited by
-
Olfactomedin 4 is a novel target gene of retinoic acids and 5-aza-2'-deoxycytidine involved in human myeloid leukemia cell growth, differentiation, and apoptosis.Blood. 2010 Dec 2;116(23):4938-47. doi: 10.1182/blood-2009-10-246439. Epub 2010 Aug 19. Blood. 2010. PMID: 20724538 Free PMC article.
-
Interferon regulatory factor-1 binds c-Cbl, enhances mitogen activated protein kinase signaling and promotes retinoic acid-induced differentiation of HL-60 human myelo-monoblastic leukemia cells.Leuk Lymphoma. 2011 Dec;52(12):2372-9. doi: 10.3109/10428194.2011.603449. Epub 2011 Aug 24. Leuk Lymphoma. 2011. PMID: 21740303 Free PMC article.
-
Resistance to discodermolide, a microtubule-stabilizing agent and senescence inducer, is 4E-BP1-dependent.Proc Natl Acad Sci U S A. 2011 Jan 4;108(1):391-6. doi: 10.1073/pnas.1016962108. Epub 2010 Dec 20. Proc Natl Acad Sci U S A. 2011. PMID: 21173253 Free PMC article.
References
-
- Tallman MS, Nabhan C, Feusner JH, Rowe JM. Acute promyelocytic leukemia: evolving therapeutic strategies. Blood. 2002;99:759–767. - PubMed
-
- Huang ME, Ye YC, hen SRC, Chai JR, Lu JX, Zhao L, Gu LJ, Wang ZY. Use of all-trans retinoic acid in the treatment of acute promyelocytic leukemia. Blood. 1988;72:567–572. - PubMed
-
- Castaigne S, Chomienne C, Daniel MT, Ballerini P, Berger R, Fenaux P, Degos L. All-trans retinoic acid as a differentiation therapy for acute promyelocytic leukemia. I. Clinical results. Blood. 1990;76:1704–1709. - PubMed
-
- Tallman MS. Differentiating therapy in acute myeloid leukemia. Leukemia. 1996;10:1262–1268. - PubMed
-
- Kambhampati S, Verma A, Li Y, Parmar S, Sassano A, Platanias LC. Signalling pathways activated by all-trans-retinoic acid in acute promyelocytic leukemia cells, Leuk. Lymphoma. 2004;45:2175–2185. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA077816/CA/NCI NIH HHS/United States
- R01 CA077816-09/CA/NCI NIH HHS/United States
- CA78282/CA/NCI NIH HHS/United States
- R01 CA078282/CA/NCI NIH HHS/United States
- CA77816/CA/NCI NIH HHS/United States
- P30 CA060553/CA/NCI NIH HHS/United States
- R01 CA100579/CA/NCI NIH HHS/United States
- R01 CA100579-02/CA/NCI NIH HHS/United States
- R01 CA121192/CA/NCI NIH HHS/United States
- CA105005/CA/NCI NIH HHS/United States
- R01 CA121192-01/CA/NCI NIH HHS/United States
- R01 CA105005/CA/NCI NIH HHS/United States
- CA100579/CA/NCI NIH HHS/United States
- CA121192/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
