

Clathrin-independent endocytosis: from nonexisting to an extreme degree of complexity
- PMID: 18193449
- PMCID: PMC2248609
- DOI: 10.1007/s00418-007-0376-5
Clathrin-independent endocytosis: from nonexisting to an extreme degree of complexity
Abstract
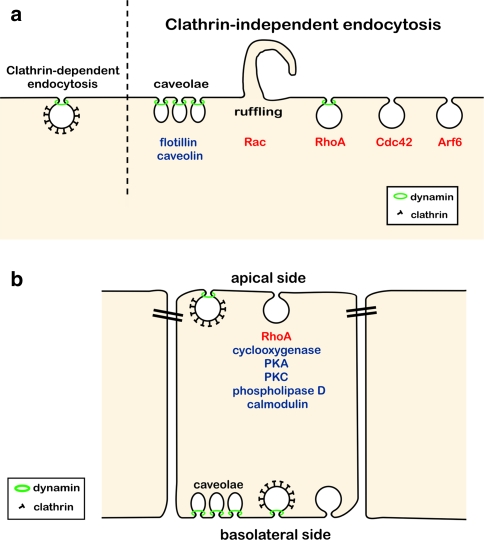
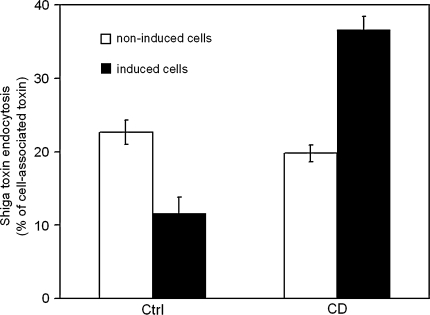
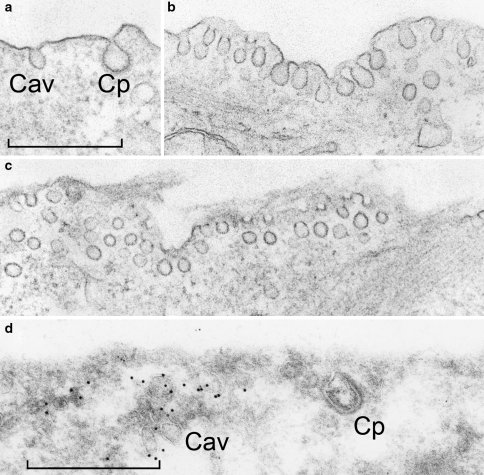
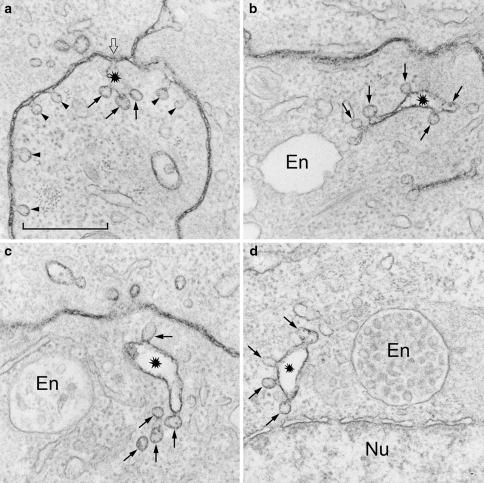
Today it is generally accepted that there are several endocytic mechanisms, both the clathrin-dependent one and mechanisms which operate without clathrin and with different requirements when it comes to dynamin, small GTP-binding proteins of the Rho family and specific lipids. It should be noted that clathrin-independent endocytosis can occur even when the cholesterol level in the membrane has been reduced to so low levels that caveolae are gone and clathrin-coated membrane areas are flat. Although new investigators in the field take it for granted that there is a multitude of entry mechanisms, it has taken a long time for this to become accepted. However, more work needs to be done, because one can still ask the question: How many endocytic mechanisms does a cell have, what are their function, and how are they regulated? This article describes some of the history of endocytosis research and attempts to give an overview of the complexity of the mechanisms and their regulation.
Figures
Similar articles
-
Porcine circovirus 2 infection of epithelial cells is clathrin-, caveolae- and dynamin-independent, actin and Rho-GTPase-mediated, and enhanced by cholesterol depletion.Virus Res. 2009 Jan;139(1):1-9. doi: 10.1016/j.virusres.2008.09.005. Epub 2008 Nov 12. Virus Res. 2009. PMID: 18952130
-
Clathrin-independent endocytosis: new insights into caveolae and non-caveolar lipid raft carriers.Biochim Biophys Acta. 2005 Dec 30;1746(3):349-63. doi: 10.1016/j.bbamcr.2005.11.007. Biochim Biophys Acta. 2005. PMID: 16440447 Review.
-
A clathrin, caveolae, and dynamin-independent endocytic pathway requiring free membrane cholesterol drives HIV-1 internalization and infection in polarized trophoblastic cells.J Mol Biol. 2007 May 18;368(5):1267-83. doi: 10.1016/j.jmb.2007.03.003. Epub 2007 Mar 12. J Mol Biol. 2007. PMID: 17395200
-
Clathrin-dependent or not: is it still the question?Traffic. 2002 Jul;3(7):443-51. doi: 10.1034/j.1600-0854.2002.30701.x. Traffic. 2002. PMID: 12047552 Review.
-
Endocytosis of gene delivery vectors: from clathrin-dependent to lipid raft-mediated endocytosis.Mol Ther. 2013 Jun;21(6):1118-30. doi: 10.1038/mt.2013.54. Epub 2013 Apr 16. Mol Ther. 2013. PMID: 23587924 Free PMC article. Review.
Cited by
-
Sorting of Clathrin-Independent Cargo Proteins Depends on Rab35 Delivered by Clathrin-Mediated Endocytosis.Traffic. 2015 Sep;16(9):994-1009. doi: 10.1111/tra.12302. Epub 2015 Jun 4. Traffic. 2015. PMID: 25988331 Free PMC article.
-
Hijacking the endocytic machinery by microbial pathogens.Protoplasma. 2010 Aug;244(1-4):75-90. doi: 10.1007/s00709-010-0164-2. Epub 2010 Jun 25. Protoplasma. 2010. PMID: 20574860 Review.
-
Compensatory endocytosis in bladder umbrella cells occurs through an integrin-regulated and RhoA- and dynamin-dependent pathway.EMBO J. 2010 Jun 16;29(12):1961-75. doi: 10.1038/emboj.2010.91. Epub 2010 May 11. EMBO J. 2010. PMID: 20461056 Free PMC article.
-
Nona-arginine facilitates delivery of quantum dots into cells via multiple pathways.J Biomed Biotechnol. 2010;2010:948543. doi: 10.1155/2010/948543. Epub 2010 Oct 27. J Biomed Biotechnol. 2010. PMID: 21048930 Free PMC article.
-
The LTB4-BLT1 axis regulates the polarized trafficking of chemoattractant GPCRs during neutrophil chemotaxis.J Cell Sci. 2018 Sep 21;131(18):jcs217422. doi: 10.1242/jcs.217422. J Cell Sci. 2018. PMID: 30158177 Free PMC article.
References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1038/ncb1627', 'is_inner': False, 'url': 'https://doi.org/10.1038/ncb1627'}, {'type': 'PubMed', 'value': '17704769', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/17704769/'}]}
- Balklava Z, Pant S, Fares H, Grant BD (2007) Genome-wide analysis identifies a general requirement for polarity proteins in endocytic traffic. Nat Cell Biol 9:1066–1073 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1111/j.1600-0854.2007.00585.x', 'is_inner': False, 'url': 'https://doi.org/10.1111/j.1600-0854.2007.00585.x'}, {'type': 'PubMed', 'value': '17547704', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/17547704/'}]}
- Benmerah A, Lamaze C (2007) Clathrin-coated pits: vive la difference? Traffic 8:970–982 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1038/ncb1260', 'is_inner': False, 'url': 'https://doi.org/10.1038/ncb1260'}, {'type': 'PubMed', 'value': '15880102', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/15880102/'}]}
- Bonazzi M, Spano S, Turacchio G, Cericola C, Valente C, Colanzi A, Kweon HS, Hsu VW, Polishchuck EV, Polishchuck RS, Sallese M, Pulvirenti T, Corda D, Luini A (2005) CtBP3/BARS drives membrane fission in dynamin-independent transport pathways. Nat Cell Biol 7:570–580 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1083/jcb.200103107', 'is_inner': False, 'url': 'https://doi.org/10.1083/jcb.200103107'}, {'type': 'PMC', 'value': 'PMC2196179', 'is_inner': False, 'url': 'https://pmc.ncbi.nlm.nih.gov/articles/PMC2196179/'}, {'type': 'PubMed', 'value': '11535619', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/11535619/'}]}
- Brown FD, Rozelle AL, Yin HL, Balla T, Donaldson JG (2001) Phosphatidylinositol 4,5-bisphosphate and Arf6-regulated membrane traffic. J Cell Biol 154:1007–1017 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
