

Receptor activator of nuclear factor kappaB ligand and osteoprotegerin regulation of bone remodeling in health and disease
- PMID: 18057140
- PMCID: PMC2528846
- DOI: 10.1210/er.2007-0014
Receptor activator of nuclear factor kappaB ligand and osteoprotegerin regulation of bone remodeling in health and disease
Abstract
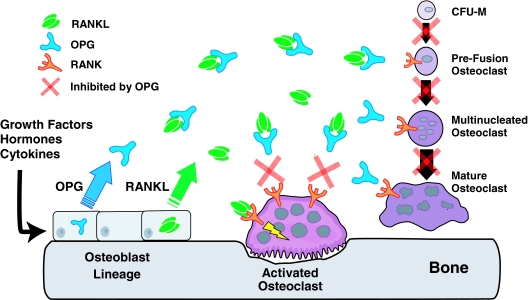
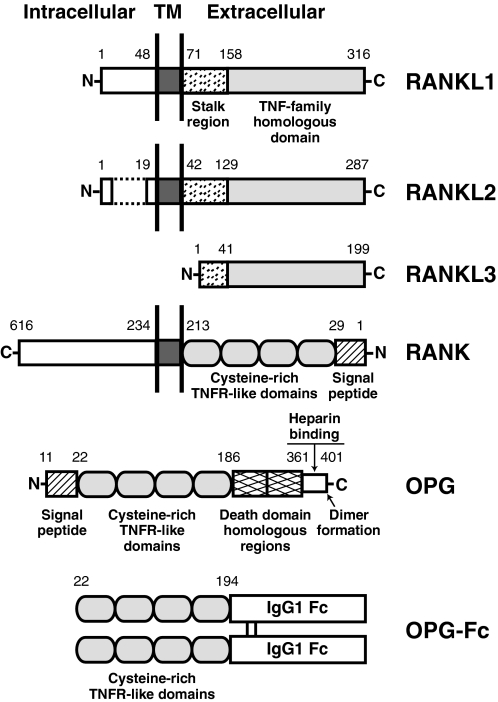
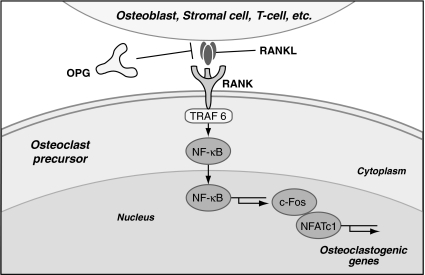
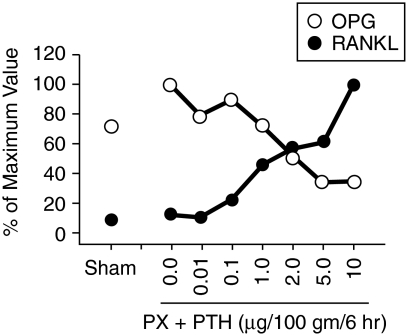
Osteoclasts and osteoblasts dictate skeletal mass, structure, and strength via their respective roles in resorbing and forming bone. Bone remodeling is a spatially coordinated lifelong process whereby old bone is removed by osteoclasts and replaced by bone-forming osteoblasts. The refilling of resorption cavities is incomplete in many pathological states, which leads to a net loss of bone mass with each remodeling cycle. Postmenopausal osteoporosis and other conditions are associated with an increased rate of bone remodeling, which leads to accelerated bone loss and increased risk of fracture. Bone resorption is dependent on a cytokine known as RANKL (receptor activator of nuclear factor kappaB ligand), a TNF family member that is essential for osteoclast formation, activity, and survival in normal and pathological states of bone remodeling. The catabolic effects of RANKL are prevented by osteoprotegerin (OPG), a TNF receptor family member that binds RANKL and thereby prevents activation of its single cognate receptor called RANK. Osteoclast activity is likely to depend, at least in part, on the relative balance of RANKL and OPG. Studies in numerous animal models of bone disease show that RANKL inhibition leads to marked suppression of bone resorption and increases in cortical and cancellous bone volume, density, and strength. RANKL inhibitors also prevent focal bone loss that occurs in animal models of rheumatoid arthritis and bone metastasis. Clinical trials are exploring the effects of denosumab, a fully human anti-RANKL antibody, on bone loss in patients with osteoporosis, bone metastasis, myeloma, and rheumatoid arthritis.
Figures
Similar articles
-
[RANKL and OPG regulation on bone remodeling].Shanghai Kou Qiang Yi Xue. 2010 Aug;19(4):443-6. Shanghai Kou Qiang Yi Xue. 2010. PMID: 20871967 Chinese.
-
CLINICAL Review #: the role of receptor activator of nuclear factor-kappaB (RANK)/RANK ligand/osteoprotegerin: clinical implications.J Clin Endocrinol Metab. 2007 Dec;92(12):4514-21. doi: 10.1210/jc.2007-0646. Epub 2007 Sep 25. J Clin Endocrinol Metab. 2007. PMID: 17895323 Review.
-
Functions of RANKL/RANK/OPG in bone modeling and remodeling.Arch Biochem Biophys. 2008 May 15;473(2):139-46. doi: 10.1016/j.abb.2008.03.018. Epub 2008 Mar 25. Arch Biochem Biophys. 2008. PMID: 18395508 Free PMC article. Review.
-
Skeletal and extraskeletal actions of denosumab.Endocrine. 2012 Aug;42(1):52-62. doi: 10.1007/s12020-012-9696-x. Epub 2012 May 13. Endocrine. 2012. PMID: 22581255 Review.
-
Key roles of the OPG-RANK-RANKL system in bone oncology.Expert Rev Anticancer Ther. 2007 Feb;7(2):221-32. doi: 10.1586/14737140.7.2.221. Expert Rev Anticancer Ther. 2007. PMID: 17288531 Review.
Cited by
-
Osteoprotegerin in bone metastases: mathematical solution to the puzzle.PLoS Comput Biol. 2012;8(10):e1002703. doi: 10.1371/journal.pcbi.1002703. Epub 2012 Oct 18. PLoS Comput Biol. 2012. PMID: 23093918 Free PMC article.
-
Effect of Blood Component Coatings of Enosseal Implants on Proliferation and Synthetic Activity of Human Osteoblasts and Cytokine Production of Peripheral Blood Mononuclear Cells.Mediators Inflamm. 2016;2016:8769347. doi: 10.1155/2016/8769347. Epub 2016 Aug 29. Mediators Inflamm. 2016. PMID: 27651560 Free PMC article.
-
Oral squamous carcinoma cells secrete RANKL directly supporting osteolytic bone loss.Oral Oncol. 2013 Feb;49(2):119-28. doi: 10.1016/j.oraloncology.2012.08.004. Epub 2012 Sep 16. Oral Oncol. 2013. PMID: 22989723 Free PMC article.
-
Assessment of efficacy of pamidronate in undifferentiated spondyloarthropathy (uSpA): a molecular mechanism.Rheumatol Int. 2013 Oct;33(10):2695. doi: 10.1007/s00296-012-2454-9. Epub 2012 Jul 26. Rheumatol Int. 2013. PMID: 22833238 No abstract available.
-
Effects of antiresorptive medications on tooth root formation and tooth eruption in paediatric patients.Orthod Craniofac Res. 2023 Dec;26 Suppl 1(Suppl 1):29-38. doi: 10.1111/ocr.12637. Epub 2023 Feb 8. Orthod Craniofac Res. 2023. PMID: 36714970 Free PMC article. Review.
References
-
- Compston JE, Vedi S, Kaptoge S, Seeman E 2007 Bone remodeling rate and remodeling balance are not co-regulated in adulthood: implications for the use of activation frequency as an index of remodeling rate. J Bone Miner Res 22:1031–1036 - PubMed
-
- Rodan GA, Martin TJ 2000 Therapeutic approaches to bone disease. Science 289:1508–1514 - PubMed
-
- Eriksen EF 1986 Normal and pathological remodeling of human trabecular bone: three dimensional reconstruction of the remodeling sequence in normals and in metabolic bone disease. Endocr Rev 7:379–408 - PubMed
-
- Simonet WS, Lacey DL, Dunstan C, Kelly M, Chang MS, Luthy R, Nhuyen HQ, Wooden S, Bennett L, Boone T, Shimamoto G, DeRose M, Elliot R, Colombero A, Tan HL, Trail G, Sullivan J, Davy E, Bucay N, Renshaw-Gegg L, Hughes TM, Hill D, Pattison W, Campbell P, Sander S, Van G, Tarpley J, Derby P, Lee R, Boyle WJ 1997 Osteoprotegerin: a novel secreted protein involved in the regulation of bone density. Cell 89:309–319 - PubMed
-
- Tsuda E, Goto M, Mochizuki SI, Yano K, Kobayashi F, Morinaga T, Higashio K 1997 Isolation of a novel cytokine from human fibroblasts that specifically inhibits osteoclastogenesis. Biochem Biophys Res Comm 234:137–142 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
