

Prominent roles for odorant receptor coding sequences in allelic exclusion
- PMID: 18045541
- PMCID: PMC2195930
- DOI: 10.1016/j.cell.2007.10.050
Prominent roles for odorant receptor coding sequences in allelic exclusion
Abstract
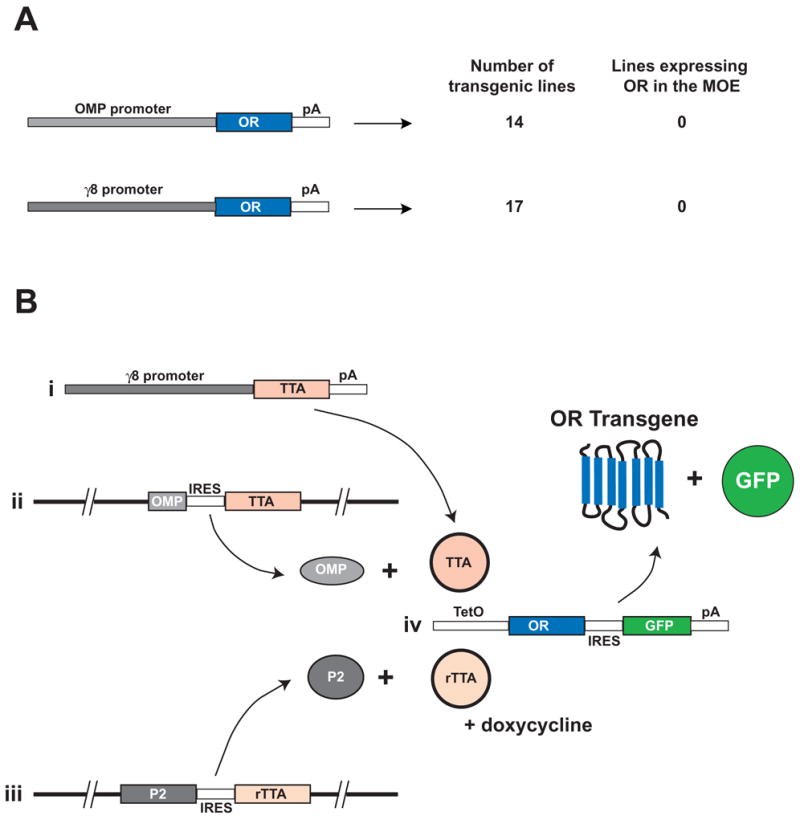
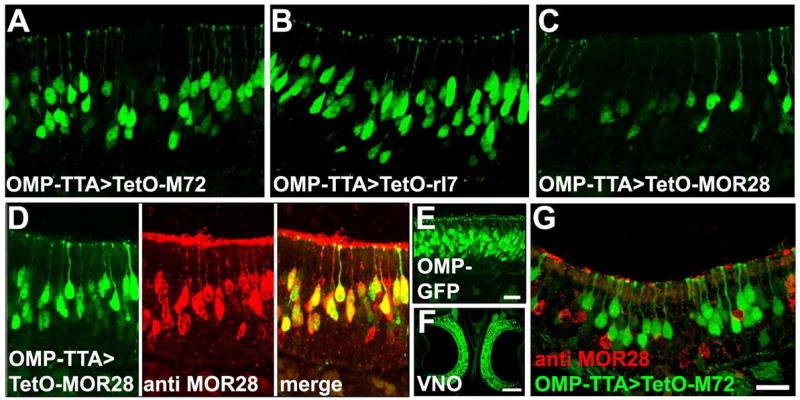
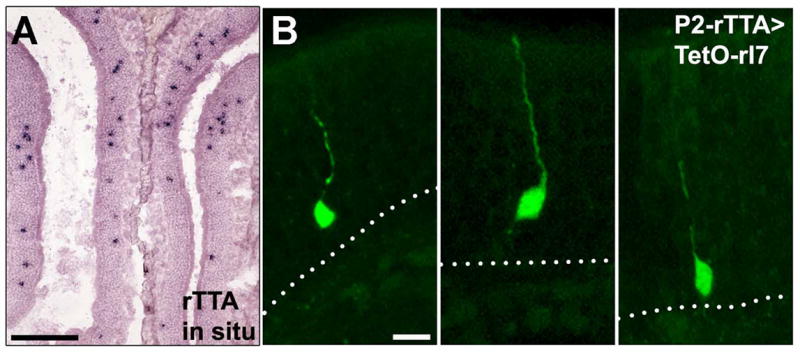
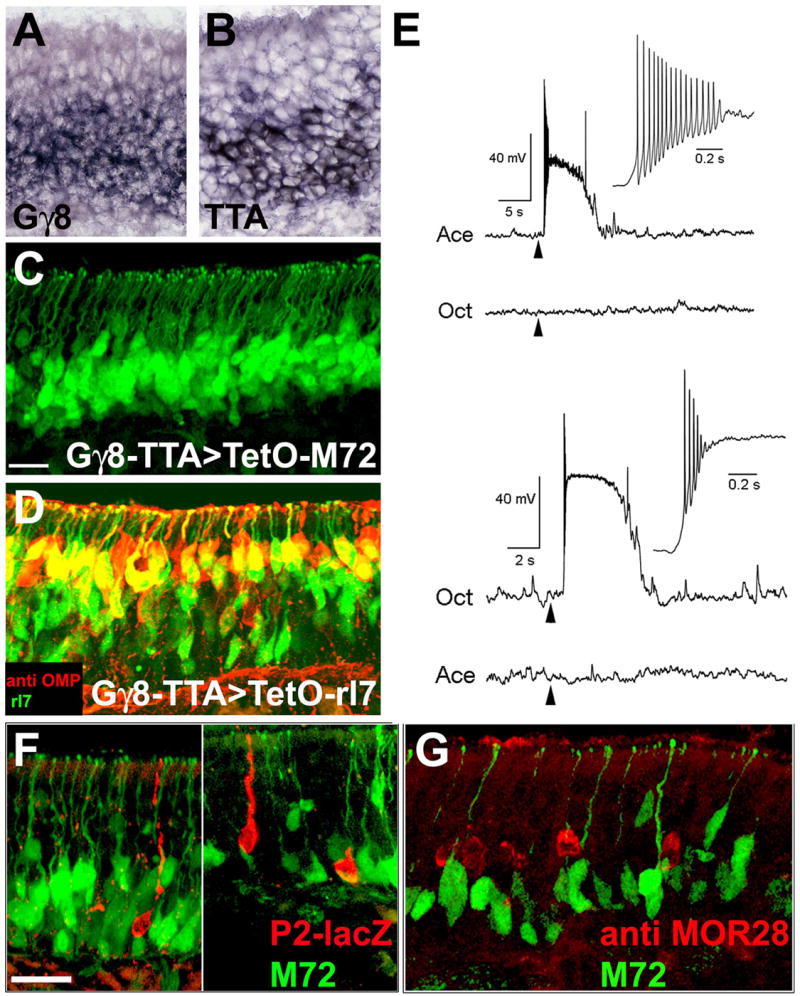
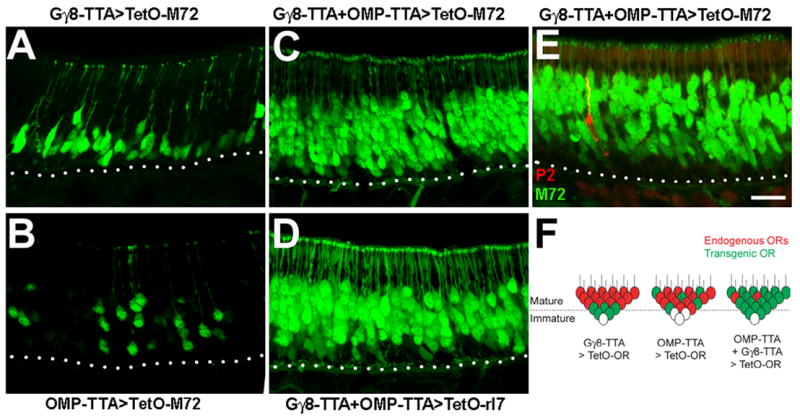
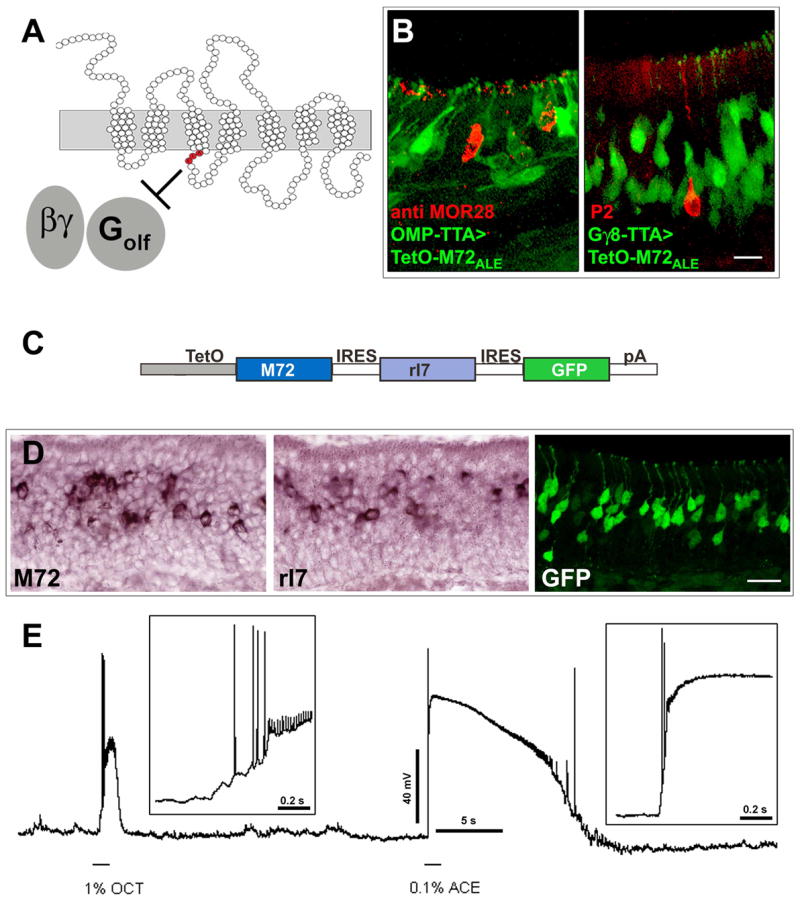
Mammalian odorant receptors (ORs) are crucial for establishing the functional organization of the olfactory system, but the mechanisms controlling their expression remain largely unexplained. Here, we utilized a transgenic approach to explore OR gene regulation. We determined that although olfactory sensory neurons (OSNs) are capable of supporting expression of multiple functional ORs, several levels of control ensure that each neuron normally expresses only a single odorant receptor. Surprisingly, this regulation extends beyond endogenous ORs even preventing expression of transgenes consisting of OR-coding sequences driven by synthetic promoters. Thus, part of the intrinsic feedback system must rely on elements present in the OR-coding sequence. Notably, by expressing the same transgenic ORs precociously in immature neurons, we have overcome this suppression and established a generic method to express any OR in approximately 90% of OSNs. These results provide important insights into the hierarchy of OR gene expression and the vital role of the OR-coding sequence in this regulation.
Figures
Comment in
-
cis-Regulatory elements within the odorant receptor coding region.Cell. 2007 Nov 30;131(5):844-6. doi: 10.1016/j.cell.2007.11.016. Cell. 2007. PMID: 18045531
Similar articles
-
Axon guidance of mouse olfactory sensory neurons by odorant receptors and the beta2 adrenergic receptor.Cell. 2004 Jun 11;117(6):833-46. doi: 10.1016/j.cell.2004.05.013. Cell. 2004. PMID: 15186782
-
Homeodomain binding motifs modulate the probability of odorant receptor gene choice in transgenic mice.Mol Cell Neurosci. 2011 Feb;46(2):381-96. doi: 10.1016/j.mcn.2010.11.001. Epub 2010 Nov 26. Mol Cell Neurosci. 2011. PMID: 21111823 Free PMC article.
-
Neuroscience. ORs rule the roost in the olfactory system.Science. 2003 Dec 19;302(5653):2078-9. doi: 10.1126/science.1093397. Science. 2003. PMID: 14684811 No abstract available.
-
One neuron-one receptor rule in the mouse olfactory system.Trends Genet. 2004 Dec;20(12):648-53. doi: 10.1016/j.tig.2004.09.006. Trends Genet. 2004. PMID: 15522461 Review.
-
Odorant receptor gene choice and axonal projection in the mouse olfactory system.Results Probl Cell Differ. 2009;47:57-75. doi: 10.1007/400_2008_3. Results Probl Cell Differ. 2009. PMID: 19083127 Review.
Cited by
-
Differentiation of developing olfactory neurons analysed in terms of coupled epigenetic landscapes.Nucleic Acids Res. 2013 May;41(9):4755-64. doi: 10.1093/nar/gkt181. Epub 2013 Mar 21. Nucleic Acids Res. 2013. PMID: 23519617 Free PMC article.
-
Laminar specific detection of APP induced neurodegeneration and recovery using MEMRI in an olfactory based Alzheimer's disease mouse model.Neuroimage. 2015 Sep;118:183-92. doi: 10.1016/j.neuroimage.2015.05.045. Epub 2015 May 27. Neuroimage. 2015. PMID: 26021215 Free PMC article.
-
An epigenetic trap stabilizes singular olfactory receptor expression.Cell. 2013 Jul 18;154(2):325-36. doi: 10.1016/j.cell.2013.06.039. Cell. 2013. PMID: 23870122 Free PMC article.
-
Functional interrogation of an odorant receptor locus reveals multiple axes of transcriptional regulation.PLoS Biol. 2013;11(5):e1001568. doi: 10.1371/journal.pbio.1001568. Epub 2013 May 21. PLoS Biol. 2013. PMID: 23700388 Free PMC article.
-
Coding odorant concentration through activation timing between the medial and lateral olfactory bulb.Cell Rep. 2012 Nov 29;2(5):1143-50. doi: 10.1016/j.celrep.2012.09.035. Epub 2012 Nov 15. Cell Rep. 2012. PMID: 23168258 Free PMC article.
References
-
- Acharya S, Karnik SS. Modulation of GDP release from transducin by the conserved Glu134-Arg135 sequence in rhodopsin. J Biol Chem. 1996;271:25406–25411. - PubMed
-
- Barnea G, O’Donnell S, Mancia F, Sun X, Nemes A, Mendelsohn M, Axel R. Odorant receptors on axon termini in the brain. Science. 2004;304:1468. - PubMed
-
- Belluscio L, Lodovichi C, Feinstein P, Mombaerts P, Katz LC. Odorant receptors instruct functional circuitry in the mouse olfactory bulb. Nature. 2002;419:296–300. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
