

Crystal structure of the yeast inner kinetochore subunit Cep3p
- PMID: 17997968
- PMCID: PMC2288795
- DOI: 10.1016/j.str.2007.09.008
Crystal structure of the yeast inner kinetochore subunit Cep3p
Abstract
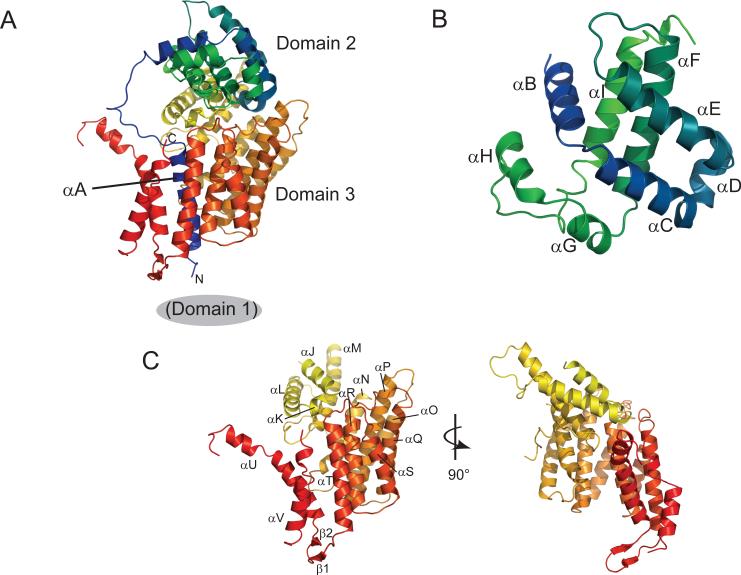
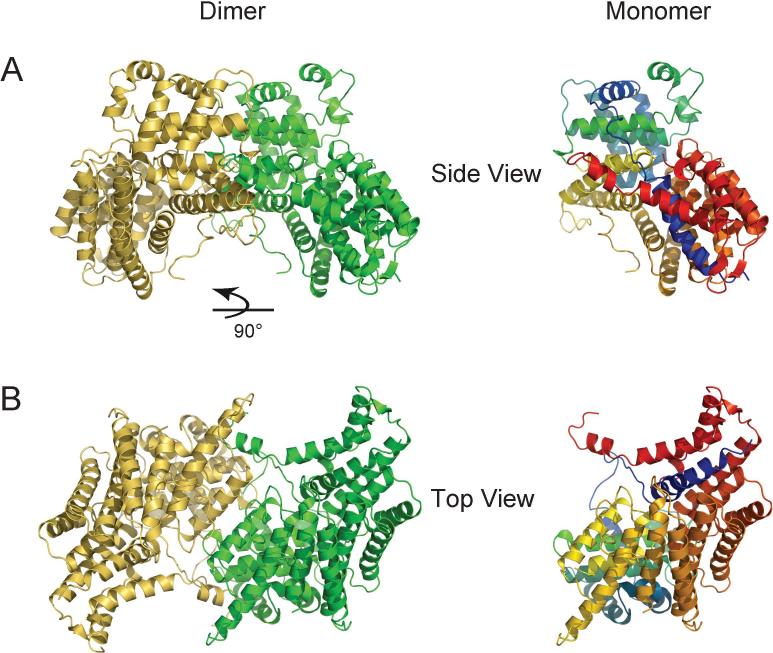
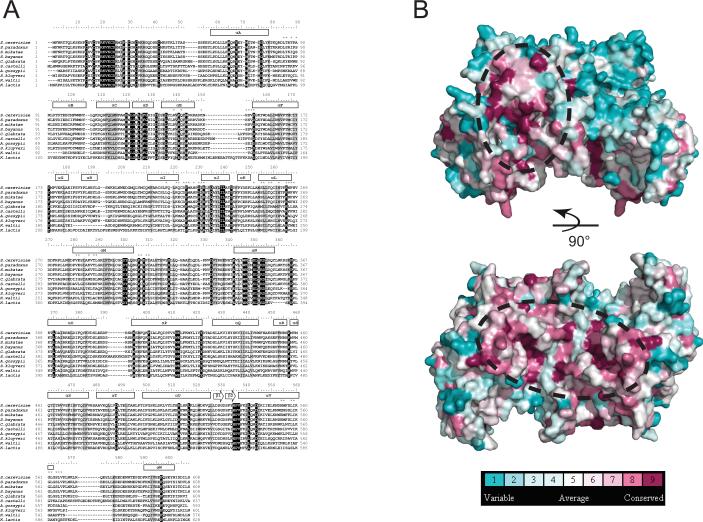
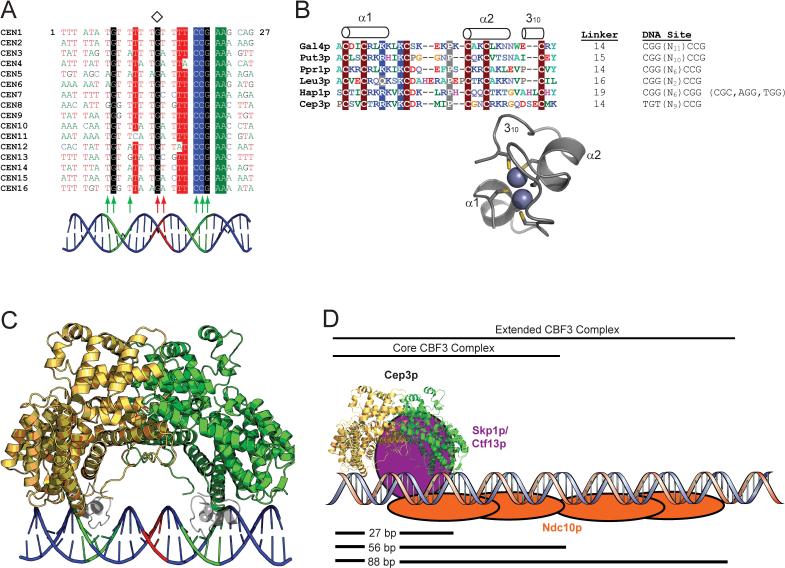
In budding yeast, the four-protein CBF3 complex (Skp1p-Ctf13p-Cep3p-Ndc10p) initiates kinetochore assembly by binding to the CDEIII locus of centromeric DNA. A Cep3p dimer recruits a Skp1p-Ctf13p heterodimer and contacts two sites on CDEIII. We report here the crystal structure, determined at 2.8 A resolution by multiple isomorphous replacement with anomalous scattering, of a truncated Cep3p (Cep3p [47-608]), comprising all but an N-terminal, Zn(2)Cys(6)-cluster, DNA-binding module. Cep3p has a well-ordered structure throughout essentially all of its polypeptide chain, unlike most yeast transcription factors, including those with Zn(2)Cys(6) clusters, such as Gal4p. This difference may reflect an underlying functional distinction: whereas any particular transcription factor must adapt to a variety of upstream activating sites, Cep3p scaffolds kinetochore assembly on centromeres uniformly configured on all 16 yeast chromosomes. We have, using the structure of Cep3p (47-608) and the known structures of Zn(2)Cys(6)-cluster domains, modeled the interaction of Cep3p with CDEIII.
Figures
Similar articles
-
Structural and functional dissection of Mif2p, a conserved DNA-binding kinetochore protein.Mol Biol Cell. 2008 Oct;19(10):4480-91. doi: 10.1091/mbc.e08-03-0297. Epub 2008 Aug 13. Mol Biol Cell. 2008. PMID: 18701705 Free PMC article.
-
Hsp90 enables Ctf13p/Skp1p to nucleate the budding yeast kinetochore.Proc Natl Acad Sci U S A. 2002 Jun 25;99(13):8585-90. doi: 10.1073/pnas.082223899. Proc Natl Acad Sci U S A. 2002. PMID: 12084919 Free PMC article.
-
Architecture of the CBF3-centromere complex of the budding yeast kinetochore.Nat Struct Mol Biol. 2018 Dec;25(12):1103-1110. doi: 10.1038/s41594-018-0154-1. Epub 2018 Nov 26. Nat Struct Mol Biol. 2018. PMID: 30478265 Free PMC article.
-
The Ndc80 complex: hub of kinetochore activity.FEBS Lett. 2007 Jun 19;581(15):2862-9. doi: 10.1016/j.febslet.2007.05.012. Epub 2007 May 11. FEBS Lett. 2007. PMID: 17521635 Review.
-
Simple centromere, complex kinetochore: linking spindle microtubules and centromeric DNA in budding yeast.J Cell Biol. 2002 Apr 15;157(2):199-203. doi: 10.1083/jcb.200201052. Epub 2002 Apr 15. J Cell Biol. 2002. PMID: 11956223 Free PMC article. Review.
Cited by
-
Structural and functional dissection of Mif2p, a conserved DNA-binding kinetochore protein.Mol Biol Cell. 2008 Oct;19(10):4480-91. doi: 10.1091/mbc.e08-03-0297. Epub 2008 Aug 13. Mol Biol Cell. 2008. PMID: 18701705 Free PMC article.
-
Molecular structures and interactions in the yeast kinetochore.Cold Spring Harb Symp Quant Biol. 2010;75:395-401. doi: 10.1101/sqb.2010.75.040. Epub 2011 Apr 5. Cold Spring Harb Symp Quant Biol. 2010. PMID: 21467141 Free PMC article.
-
Structural basis for assembly of the CBF3 kinetochore complex.EMBO J. 2018 Jan 17;37(2):269-281. doi: 10.15252/embj.201798134. Epub 2017 Dec 6. EMBO J. 2018. PMID: 29212814 Free PMC article.
-
Structure of yeast kinetochore Ndc10 DNA-binding domain reveals unexpected evolutionary relationship to tyrosine recombinases.J Biol Chem. 2012 Feb 10;287(7):5173-9. doi: 10.1074/jbc.C111.318501. Epub 2012 Jan 3. J Biol Chem. 2012. PMID: 22215672 Free PMC article.
-
Ndc10 is a platform for inner kinetochore assembly in budding yeast.Nat Struct Mol Biol. 2011 Dec 4;19(1):48-55. doi: 10.1038/nsmb.2178. Nat Struct Mol Biol. 2011. PMID: 22139014 Free PMC article.
References
-
- Abrahams JP, Leslie AG. Methods used in the structure determination of bovine mitochondrial F1 ATPase. Acta Crystallogr D Biol Crystallogr. 1996;52:30–42. - PubMed
-
- Bricogne G, Vonrhein C, Flensburg C, Schiltz M, Paciorek W. Generation, representation and flow of phase information in structure determination: recent developments in and around SHARP 2.0. Acta Crystallogr D Biol Crystallogr. 2003;59:2023–2030. - PubMed
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr. 1998;54:905–921. - PubMed
-
- Camahort R, Li B, Florens L, Swanson SK, Washburn MP, Gerton JL. Scm3 is essential to recruit the histone h3 variant cse4 to centromeres and to maintain a functional kinetochore. Mol Cell. 2007;26:853–865. - PubMed
-
- Choo KH. Domain organization at the centromere and neocentromere. Dev Cell. 2001;1:165–177. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
