

An interlocked dimer of the protelomerase TelK distorts DNA structure for the formation of hairpin telomeres
- PMID: 17889664
- PMCID: PMC2041798
- DOI: 10.1016/j.molcel.2007.07.026
An interlocked dimer of the protelomerase TelK distorts DNA structure for the formation of hairpin telomeres
Abstract
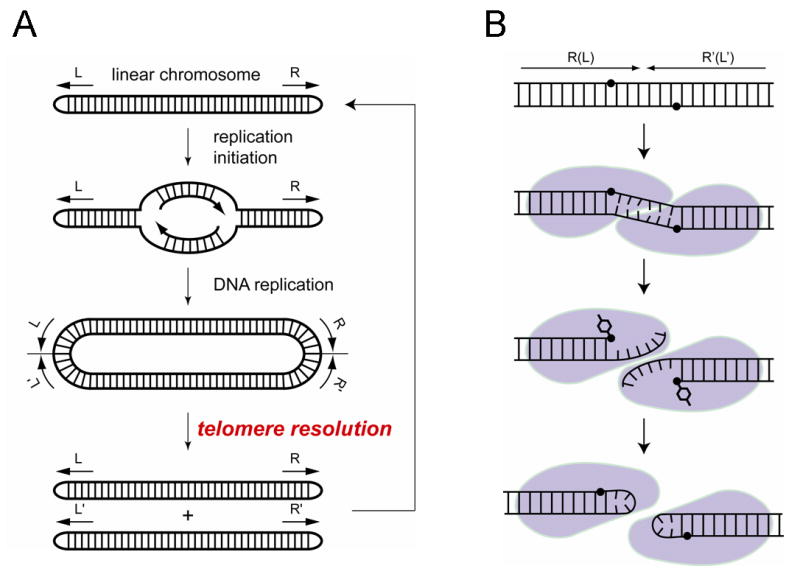
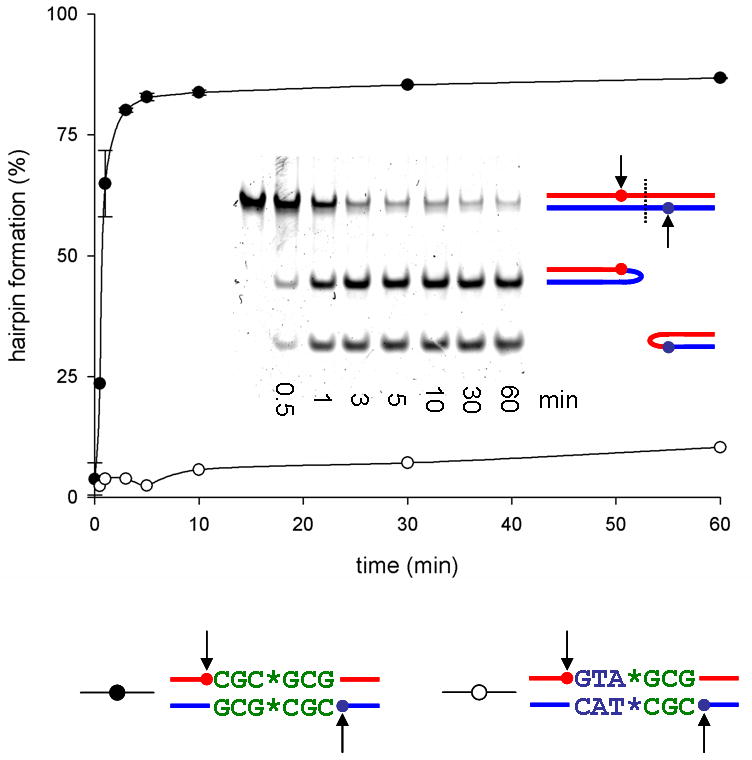
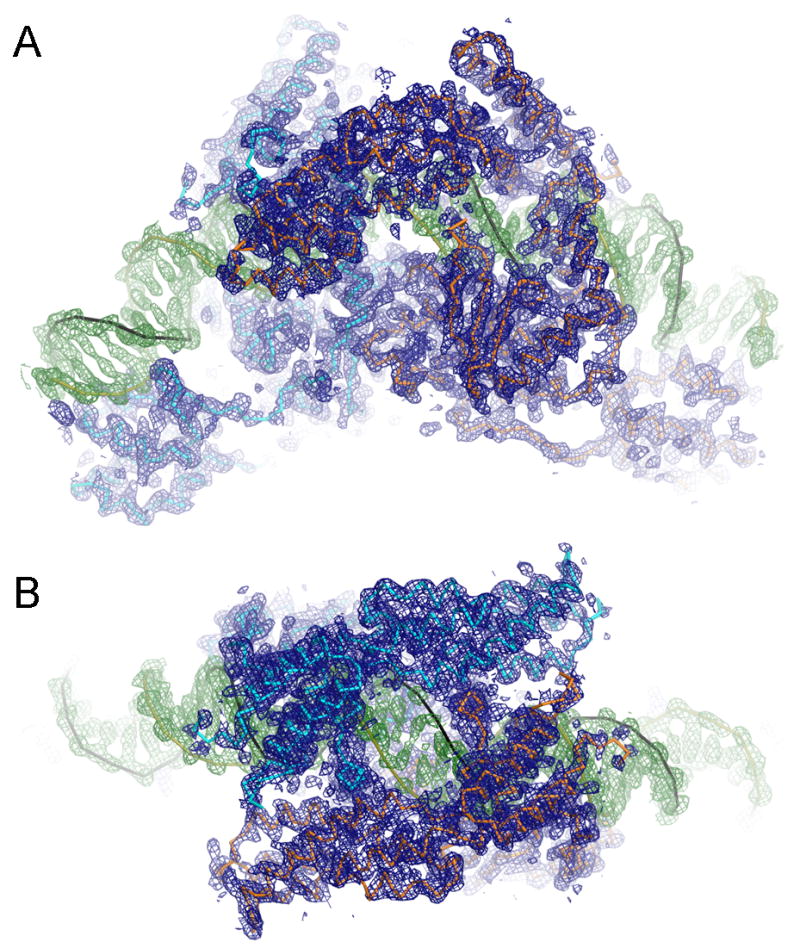
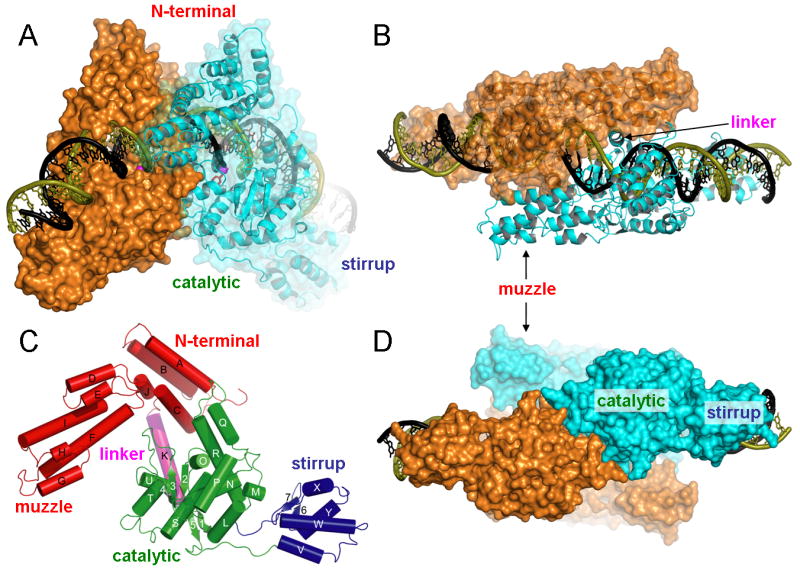
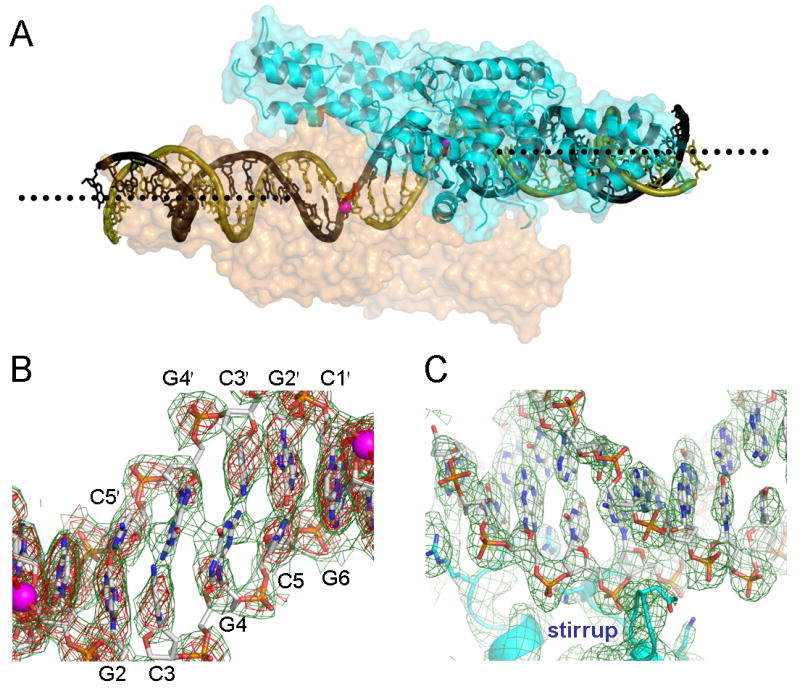
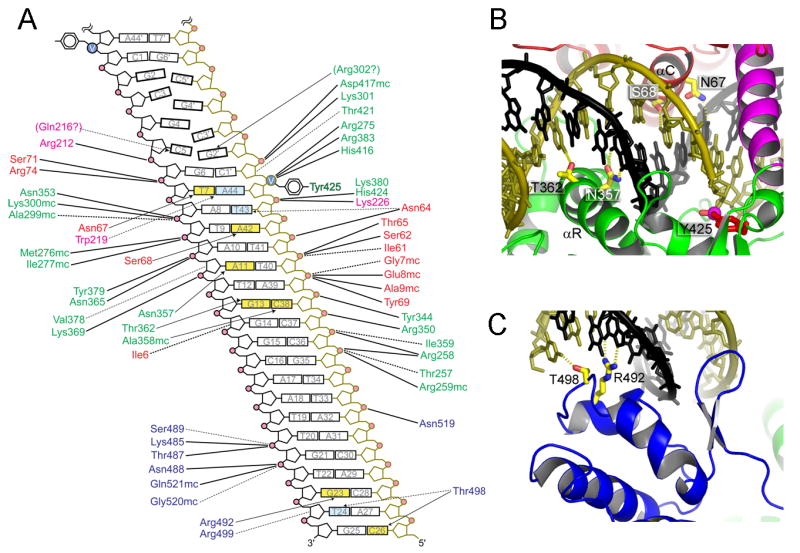
The termini of linear chromosomes are protected by specialized DNA structures known as telomeres that also facilitate the complete replication of DNA ends. The simplest type of telomere is a covalently closed DNA hairpin structure found in linear chromosomes of prokaryotes and viruses. Bidirectional replication of a chromosome with hairpin telomeres produces a catenated circular dimer that is subsequently resolved into unit-length chromosomes by a dedicated DNA cleavage-rejoining enzyme known as a hairpin telomere resolvase (protelomerase). Here we report a crystal structure of the protelomerase TelK from Klebsiella oxytoca phage varphiKO2, in complex with the palindromic target DNA. The structure shows the TelK dimer destabilizes base pairing interactions to promote the refolding of cleaved DNA ends into two hairpin ends. We propose that the hairpinning reaction is made effectively irreversible by a unique protein-induced distortion of the DNA substrate that prevents religation of the cleaved DNA substrate.
Figures

Similar articles
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Comparison of Two Modern Survival Prediction Tools, SORG-MLA and METSSS, in Patients With Symptomatic Long-bone Metastases Who Underwent Local Treatment With Surgery Followed by Radiotherapy and With Radiotherapy Alone.Clin Orthop Relat Res. 2024 Dec 1;482(12):2193-2208. doi: 10.1097/CORR.0000000000003185. Epub 2024 Jul 23. Clin Orthop Relat Res. 2024. PMID: 39051924
-
Qualitative evidence synthesis informing our understanding of people's perceptions and experiences of targeted digital communication.Cochrane Database Syst Rev. 2019 Oct 23;10(10):ED000141. doi: 10.1002/14651858.ED000141. Cochrane Database Syst Rev. 2019. PMID: 31643081 Free PMC article.
-
A Blog-Based Study of Autistic Adults' Experiences of Aloneness and Connection and the Interplay with Well-Being: Corpus-Based and Thematic Analyses.Autism Adulthood. 2023 Dec 1;5(4):437-449. doi: 10.1089/aut.2022.0073. Epub 2023 Dec 12. Autism Adulthood. 2023. PMID: 38116056 Free PMC article.
-
Trends in Surgical and Nonsurgical Aesthetic Procedures: A 14-Year Analysis of the International Society of Aesthetic Plastic Surgery-ISAPS.Aesthetic Plast Surg. 2024 Oct;48(20):4217-4227. doi: 10.1007/s00266-024-04260-2. Epub 2024 Aug 5. Aesthetic Plast Surg. 2024. PMID: 39103642 Review.
Cited by
-
ALOG domains: provenance of plant homeotic and developmental regulators from the DNA-binding domain of a novel class of DIRS1-type retroposons.Biol Direct. 2012 Nov 12;7:39. doi: 10.1186/1745-6150-7-39. Biol Direct. 2012. PMID: 23146749 Free PMC article.
-
The telomere resolvase, TelA, utilizes an underwound pre-cleavage intermediate to promote hairpin telomere formation.PLoS One. 2023 Nov 29;18(11):e0294732. doi: 10.1371/journal.pone.0294732. eCollection 2023. PLoS One. 2023. PMID: 38019799 Free PMC article.
-
Preventing broken Borrelia telomeres: ResT couples dual hairpin telomere formation with product release.J Biol Chem. 2010 Dec 24;285(52):41010-8. doi: 10.1074/jbc.M110.150060. Epub 2010 Oct 14. J Biol Chem. 2010. PMID: 20952394 Free PMC article.
-
Structure analysis of the telomere resolvase from the Lyme disease spirochete Borrelia garinii reveals functional divergence of its C-terminal domain.Nucleic Acids Res. 2024 Aug 12;52(14):8431-8442. doi: 10.1093/nar/gkae580. Nucleic Acids Res. 2024. PMID: 38979576 Free PMC article.
-
Spring loading a pre-cleavage intermediate for hairpin telomere formation.Nucleic Acids Res. 2015 Jul 13;43(12):6062-74. doi: 10.1093/nar/gkv497. Epub 2015 May 24. Nucleic Acids Res. 2015. PMID: 26007659 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
