

doi: 10.1038/nsmb1300.
Epub 2007 Sep 16.
The structure of bacterial ParM filaments
Affiliations
- PMID: 17873883
- PMCID: PMC3541950
- DOI: 10.1038/nsmb1300
Item in Clipboard
The structure of bacterial ParM filaments
Nat Struct Mol Biol.
2007 Oct.
Abstract
Bacterial ParM is a homolog of eukaryotic actin and is involved in moving plasmids so that they segregate properly during cell division. Using cryo-EM and three-dimensional reconstruction, we show that ParM filaments have a different structure from F-actin, with very different subunit-subunit interfaces. These interfaces result in the helical handedness of the ParM filament being opposite to that of F-actin. Like F-actin, ParM filaments have a variable twist, and we show that this involves domain-domain rotations within the ParM subunit. The present results yield new insights into polymorphisms within F-actin, as well as the evolution of polymer families.
Figures

Electron micrographs of ParM. (a) Negatively stained ParM filaments. Scale bars, 1,000 Å. (b) Cryo-EM of unstained frozen-hydrated ParM filaments on a carbon substrate. (c) A power spectrum computed from 3,453 nonoverlapping cryo-EM segments of ParM filaments, where each segment was 480 Å in length. Four layer lines are labeled with their Bessel orders. The n = −2 line (arising from a left-handed two-start helix) is at ~1/(307 Å), the n = +3 line (arising from a right-handed three-start helix) is at ~1/(65 Å), the n +1 line (from a right-handed one-start helix) is at 1/(53.4 Å), and the n = −1 line (from a left-handed one-start helix) is at 1/(45.5 Å). (d) Quick-freeze/deep-etch electron micrograph of ParM filaments. Such images confirmed that the long-pitch helices repeating every ~300 Å are left-handed, not right-handed as in F-actin. As a control, F-actin filaments were imaged both separately from ParM and after preparation with ParM. (e,f) Power spectra computed from quick-freeze/deep-etch images of ParM (e) and F-actin (f) clearly show the reversal of helical handedness. Vertical lines in e,f are the meridians of the power spectra. White arrows indicate the reflections arising from the long-pitch helices, left-handed at B1/(300 Å) in ParM (e) and right-handed at ~1/(360 Å) in F-actin (f). Gray arrows indicate the reflections from the one-start helix, right-handed in ParM at ~1/(53 Å) (e) and left-handed in F-actin at B1/(59 Å) (f). (g,h) The quick-freeze/deep-etch images can be averaged, and this is shown for ParM (g) and F-actin (h). The left-handed long-pitch helix (repeating every ~300 Å) in ParM (g) and the left-handed short-pitch helix in F-actin (repeating every 59 Å) (h) are indicated with black lines.
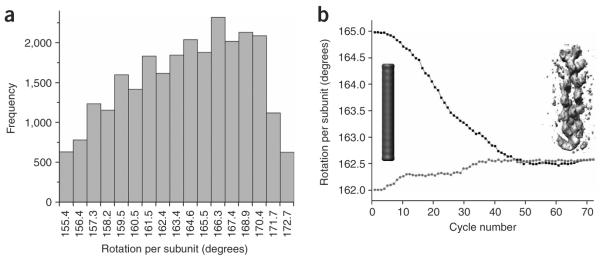
Variable twist, and overcoming it with the IHRSR approach. (a) Histogram of average twists for segments of frozen-hydrated ParM filaments. (b) The convergence of the IHRSR procedure is shown for cryo-EM segments of ParM classified as having a mean twist of ~162.5°. A solid featureless object (cylinder, left) was used as an initial reference. The IHRSR procedure was run twice, starting the search at either 162.0° (●) or 165.0° (■). After ~50 cycles, the two different sets converged to the same symmetry and the same reconstructed volume. An unsymmetrized volume from the seventieth cycle is shown as gray density at right. Each cycle, such a volume is analyzed to determine the symmetry within it, and this symmetry is imposed on the volume to generate a new symmetrized reference for the next cycle. The unsymmetrized volume shown has not been corrected for the CTF.
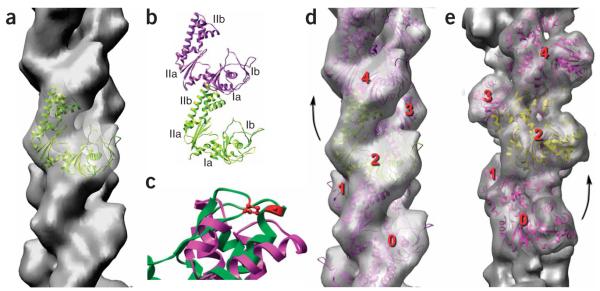
Filaments of ParM have the opposite handedness to that of F-actin. (a–d) The cryo-EM reconstruction of ParM is shown as a transparent gray surface in a and d, and a model for the ParM subunit (ribbon representation) has been fit into this volume. The four subdomains of the ParM subunit are labeled in b; the strongest interface in the ParM filament is between subdomain IIa of one subunit and subdomain Ia of a subunit above it. c shows a close-up view of an alignment of ParM subdomain IIa (magenta) with actin subdomain 4 (green, with actin residues 204 and 247 in red). Actin residues 204 and 247 are expected to be part of the subunit-subunit interface in F-actin, as mutation of these residues results in loss of polymerization. The overall structural conservation between actin and ParM does not extend to this region, consistent with different filamentous interfaces. (e) For comparison, a reconstruction of F-actin is shown. In F-actin (e), subunit 2 is rotated from subunit 0 by a right-handed rotation that is ~28° on average. In ParM (d), subunit 4 is rotated from subunit 2 by a left-handed rotation that is ~29° on average.
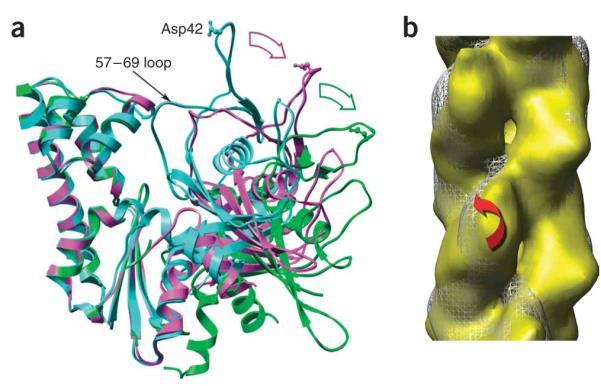
Domain rotations within ParM. (a) Crystal structures of ParM in the closed state (PDB 1MWM, cyan) and the open state (PDB 1MWK, magenta) are compared with the subunit model (green) used to fit the EM reconstruction (Fig. 3a,b). Major domain II has been aligned for all three structures. The 57–69 loop in domain I is one of the few elements that shifts between the crystal structures of the open and closed states. A single residue, Asp42, is shown in ball-and-stick representation to indicate the relative motions of domain I. The displacement of Asp42 from 1MWM to 1MWK is ~18 Å, which is about the same as the displacement from 1MWK to the EM model. (b) Reconstructions of ParM in two different twist states are compared. White mesh, twist of 165.2°; yellow surface, twist of 169.6°. Red arrow represents rotation of major domain I by ~10° between these two reconstructions while the orientation of major domain II remains relatively fixed.
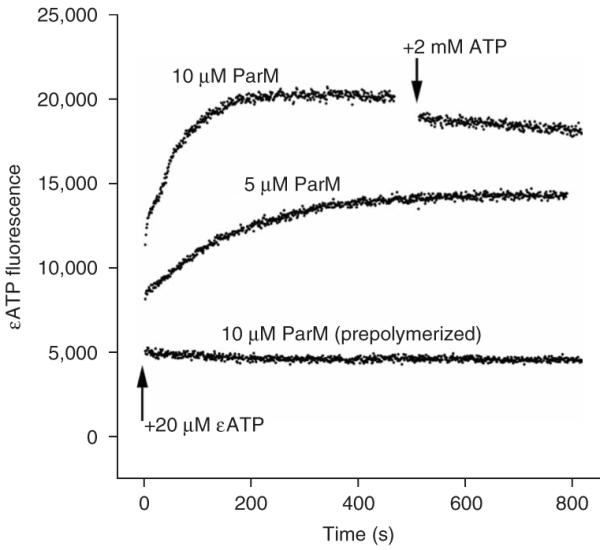
Absence of nucleotide exchange after ParM polymerization. Fluorescent etheno-ATP (εATP) was used to induce polymerization in the nonhydrolyzing ParM mutant E148A. Acrylamide was used as a collisional quencher to increase the signal from the bound nucleotide and to monitor protection of the nucleotide from interaction with solvent. Adding ParM (5 μM) to εATP caused a rapid jump in fluorescence, consistent with monomer binding, followed by a slower increase, whose time course is identical to that of polymerization. When prepolymerized ParM (10 μM) was added to εATP, there was no jump in fluorescence. When a 100-fold excess of unlabeled ATP was added to the polymerized ParM, only a very slow decrease in fluorescence (with a half-life on the order of 1 h) was seen that was not consistent with rapid nucleotide exchange on the filament. This time course is orders of magnitude slower than exchange on monomeric ParM, and the kinetics are consistent with turnover of subunits in the filament.
Similar articles
-
Structures of actin-like ParM filaments show architecture of plasmid-segregating spindles.Nature. 2015 Jul 2;523(7558):106-10. doi: 10.1038/nature14356. Epub 2015 Apr 27. Nature. 2015. PMID: 25915019 Free PMC article.
-
Electron cryomicroscopy of E. coli reveals filament bundles involved in plasmid DNA segregation.Science. 2009 Jan 23;323(5913):509-12. doi: 10.1126/science.1164346. Epub 2008 Dec 18. Science. 2009. PMID: 19095899
-
F-actin-like filaments formed by plasmid segregation protein ParM.EMBO J. 2002 Dec 16;21(24):6935-43. doi: 10.1093/emboj/cdf672. EMBO J. 2002. PMID: 12486014 Free PMC article.
-
A tale of two polymers: new insights into helical filaments.Nat Rev Mol Cell Biol. 2003 Aug;4(8):621-30. doi: 10.1038/nrm1176. Nat Rev Mol Cell Biol. 2003. PMID: 12923524 Review.
-
Actin's prokaryotic homologs.Curr Opin Struct Biol. 2003 Apr;13(2):244-8. doi: 10.1016/s0959-440x(03)00027-7. Curr Opin Struct Biol. 2003. PMID: 12727519 Review.
Cited by
-
Archaeal actin from a hyperthermophile forms a single-stranded filament.Proc Natl Acad Sci U S A. 2015 Jul 28;112(30):9340-5. doi: 10.1073/pnas.1509069112. Epub 2015 Jun 29. Proc Natl Acad Sci U S A. 2015. PMID: 26124094 Free PMC article.
-
Structural plasticity in actin and tubulin polymer dynamics.Science. 2009 Aug 21;325(5943):960-3. doi: 10.1126/science.1168823. Science. 2009. PMID: 19696342 Free PMC article. Review.
-
When cytoskeletal worlds collide.Proc Natl Acad Sci U S A. 2010 Nov 16;107(46):19609-10. doi: 10.1073/pnas.1014665107. Epub 2010 Nov 8. Proc Natl Acad Sci U S A. 2010. PMID: 21059902 Free PMC article. No abstract available.
-
Phylogenetic analysis identifies many uncharacterized actin-like proteins (Alps) in bacteria: regulated polymerization, dynamic instability and treadmilling in Alp7A.Mol Microbiol. 2009 Aug;73(4):534-52. doi: 10.1111/j.1365-2958.2009.06771.x. Epub 2009 Jul 7. Mol Microbiol. 2009. PMID: 19602153 Free PMC article.
-
ParA2, a Vibrio cholerae chromosome partitioning protein, forms left-handed helical filaments on DNA.Proc Natl Acad Sci U S A. 2010 Mar 9;107(10):4590-5. doi: 10.1073/pnas.0913060107. Epub 2010 Feb 22. Proc Natl Acad Sci U S A. 2010. PMID: 20176965 Free PMC article.
References
-
- van den Ent F, Amos LA, Löwe J. Prokaryotic origin of the actin cytoskeleton. Nature. 2001;413:39–44. - PubMed
-
- Lowe J, Amos LA. Crystal structure of the bacterial cell-division protein FtsZ. Nature. 1998;391:203–206. - PubMed
-
- Egelman EH. A robust algorithm for the reconstruction of helical filaments using single-particle methods. Ultramicroscopy. 2000;85:225–234. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
