

doi: 10.1128/JVI.01517-07.
Epub 2007 Sep 12.
DDX3 DEAD-box RNA helicase is required for hepatitis C virus RNA replication
Affiliations
- PMID: 17855521
- PMCID: PMC2168844
- DOI: 10.1128/JVI.01517-07
Item in Clipboard
DDX3 DEAD-box RNA helicase is required for hepatitis C virus RNA replication
J Virol.
2007 Dec.
Abstract
DDX3, a DEAD-box RNA helicase, binds to the hepatitis C virus (HCV) core protein. However, the role(s) of DDX3 in HCV replication is still not understood. Here we demonstrate that the accumulation of both genome-length HCV RNA (HCV-O, genotype 1b) and its replicon RNA were significantly suppressed in HuH-7-derived cells expressing short hairpin RNA targeted to DDX3 by lentivirus vector transduction. As well, RNA replication of JFH1 (genotype 2a) and release of the core into the culture supernatants were suppressed in DDX3 knockdown cells after inoculation of the cell culture-generated HCVcc. Thus, DDX3 is required for HCV RNA replication.
Figures
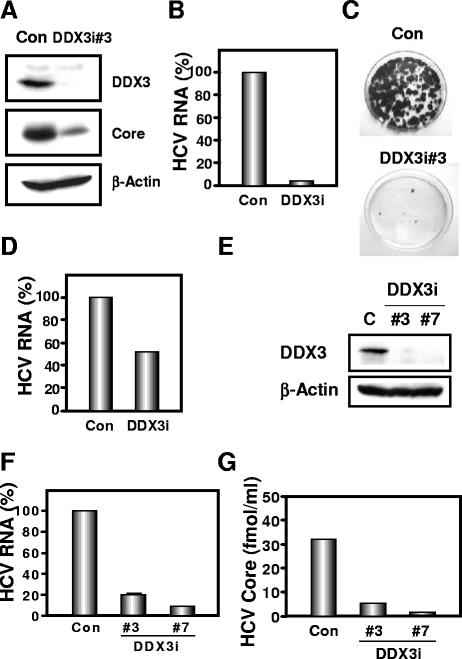
Requirement of DDX3 for HCV replication. (A to D) Effect of DDX3 knockdown on HCV RNA replication. (A) Inhibition of DDX3 expression by shRNA-producing lentivirus vector. The results of Western blot analysis of cellular lysates with anti-DDX3 (ProSci), anti-HCV core (CP-9; Institute of Immunology), or an anti-β-actin antibody (Sigma) in O cells expressing shRNA against DDX3 (DDX3i#3) as well as in O cells transduced with a control lentivirus vector (Con) are shown. (B) The level of genome-length HCV RNA was monitored by real-time LightCycler PCR (Roche). Experiments were done in duplicate, and bars represent the mean percentages of HCV RNA. (C) Efficiency of colony formation in DDX3 knockdown cells. In vitro-transcribed ON/C-5B K1609E RNA (2 μg) was transfected into the DDX3 knockdown Oc cells (DDX3i#3) or the Oc cells transduced with a control lentivirus vector (Con). G418-resistant colonies were stained with Coomassie brilliant blue at 3 weeks after electroporation of RNA. Experiments were done in duplicate, and representative results are shown. (D) The level of subgenomic replicon RNA was monitored by real-time LightCycler PCR. Experiments were done in duplicate, and bars represent the mean percentages of HCV RNA. (E to G) Effect of DDX3 knockdown on HCV infection. (E) Inhibition of DDX3 expression by shRNA-producing lentivirus vector. The results of Western blot analysis of cellular lysates with anti-DDX3 or an anti-β-actin antibody for RSc cells expressing the shRNA DDX3i#3 or DDX3i#7 and for RSc cells transduced with a control lentivirus vector (Con) are shown. (F) The level of genome-length HCV (JFH1) RNA was monitored by real-time LightCycler PCR after inoculation of the cell culture-generated HCVcc. Results from three independent experiments are shown. (G) The levels of the HCV core in the culture supernatants were determined by an enzyme-linked immunosorbent assay (Mitsubishi Kagaku Bio-Clinical Laboratories). Experiments were done in duplicate, and bars represent the mean HCV core protein levels.
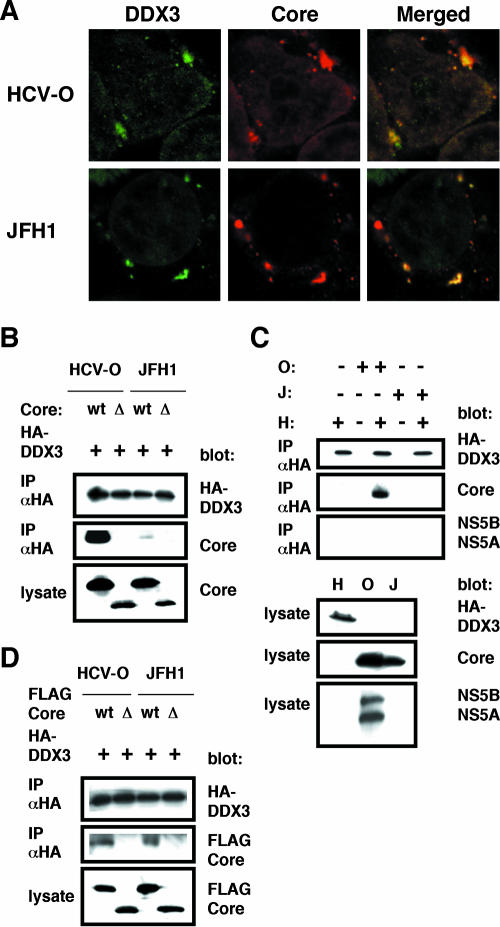
Interaction of the HCV core with DDX3. (A) The HCV core colocalizes with DDX3. 293FT cells cotransfected with 100 ng of pCXbsr/core(HCV-O) or pcDNA3/core(JFH1) and 200 ng of pHA-DDX3 were examined by confocal laser scanning microscopy. Cells were stained with anti-HCV core (CP-9 and CP-11 mixture) and anti-DDX3 antibodies and were then visualized with fluorescein isothiocyanate (DDX3) or Cy3 (core). Images were visualized using confocal laser scanning microscopy (LSM510; Carl Zeiss). The right panels exhibit the two-color overlay images (Merged). Colocalization is shown in yellow. (B) The core binds to DDX3. 293FT cells were cotransfected with 4 μg of pHA-DDX3 and 4 μg of pCXbsr/core(HCV-O) (wt), pcDNA3/Δcore(HCV-O) (Δ), pcDNA3/core(JFH1) (wt), or pcDNA3/Δcore(JFH1) (Δ). The cell lysates were immunoprecipitated with an anti-HA antibody (3F10; Roche), followed by immunoblot analysis using either anti-HA (HA-7; Sigma) or anti-HCV core antibody (CP-9 and CP-11 mixture). (C) 293FT cells transfected with 4 μg of pHA-DDX3 (H), O cells (O), or RSc cells 3 days after inoculation of HCVcc (JFH1) (J) were lysed and immunoprecipitated (IP) with 1 μg of anti-HA antibody (3F10), followed by immunoblotting with anti-HA (HA-7), anti-core (CP-9 and CP-11 mixture), or anti-HCV NS5A (no. 8926) and anti-HCV NS5B. (D) 293FT cells transfected with 4 μg of pHA-DDX3 and 4 μg of pcDNA3/FLAG-core(HCV-O) (wt), pcDNA3/FLAG-Δcore(HCV-O) (Δ), pcDNA3/FLAG-core(JFH1) (wt), or pcDNA3/FLAG-Δcore(JFH1) (Δ) were lysed and immunoprecipitated with 1 μg of an anti-HA antibody (3F10), followed by immunoblotting with an anti-HA (HA-7) or anti-core (CP-9 and CP-11 mixture) antibody.
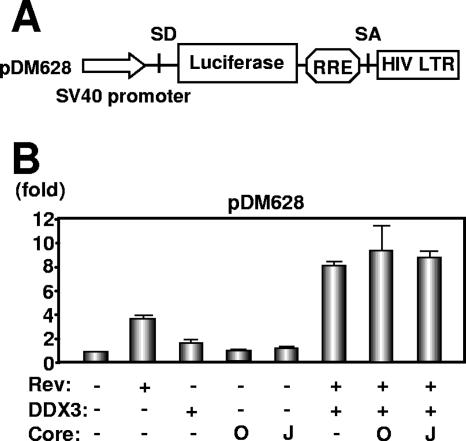
HCV core does not affect the DDX3-mediated synergistic activation of Rev function. (A) Schematic representation of HIV-1 Rev-dependent luciferase-based reporter plasmid pDM628 harboring a splicing donor (SD), splicing acceptor (SA), and RRE. (B) 293FT cells were cotransfected with 100 ng of pDM628, 200 ng of pcRev, 200 ng of pHA-DDX3, and/or 100 ng of either pcDNA3/core(HCV-O) (O) or pcDNA3/core(JFH1) (J). A luciferase assay was performed 24 h later. All transfections utilized equal total amounts of plasmid DNA owing to the addition of the empty vector pcDNA3 to the transfection mixture. The relative stimulation of luciferase activity (n-fold) is shown. The results shown are means from three independent experiments.
Similar articles
-
Hepatitis C virus core protein interacts with a human DEAD box protein DDX3.Virology. 1999 May 10;257(2):330-40. doi: 10.1006/viro.1999.9659. Virology. 1999. PMID: 10329544
-
DDX3, a DEAD box RNA helicase, is deregulated in hepatitis virus-associated hepatocellular carcinoma and is involved in cell growth control.Oncogene. 2006 Mar 30;25(14):1991-2003. doi: 10.1038/sj.onc.1209239. Oncogene. 2006. PMID: 16301996
-
Cell culture-adaptive NS3 mutations required for the robust replication of genome-length hepatitis C virus RNA.Virus Res. 2007 Apr;125(1):88-97. doi: 10.1016/j.virusres.2006.12.011. Epub 2007 Jan 18. Virus Res. 2007. PMID: 17239465
-
[Replication of hepatitis C virus genome].Uirusu. 2008 Dec;58(2):191-8. Uirusu. 2008. PMID: 19374197 Review. Japanese.
-
In vitro replication models for the hepatitis C virus.J Viral Hepat. 2007 Jan;14(1):2-10. doi: 10.1111/j.1365-2893.2006.00807.x. J Viral Hepat. 2007. PMID: 17212638 Review.
Cited by
-
EWSR1 binds the hepatitis C virus cis-acting replication element and is required for efficient viral replication.J Virol. 2013 Jun;87(12):6625-34. doi: 10.1128/JVI.01006-12. Epub 2013 Apr 3. J Virol. 2013. PMID: 23552423 Free PMC article.
-
Caspase-Dependent Cleavage of DDX21 Suppresses Host Innate Immunity.mBio. 2021 Jun 29;12(3):e0100521. doi: 10.1128/mBio.01005-21. Epub 2021 Jun 14. mBio. 2021. PMID: 34125604 Free PMC article.
-
Cytoplasmic dsRNA induces the expression of OCT3/4 and NANOG mRNAs in differentiated human cells.J Biol Chem. 2019 Dec 13;294(50):18969-18979. doi: 10.1074/jbc.RA119.009783. Epub 2019 Oct 15. J Biol Chem. 2019. PMID: 31615841 Free PMC article.
-
P bodies, stress granules, and viral life cycles.Cell Host Microbe. 2008 Apr 17;3(4):206-12. doi: 10.1016/j.chom.2008.03.004. Cell Host Microbe. 2008. PMID: 18407064 Free PMC article. Review.
-
Viral targeting of DEAD box protein 3 reveals its role in TBK1/IKKepsilon-mediated IRF activation.EMBO J. 2008 Aug 6;27(15):2147-57. doi: 10.1038/emboj.2008.143. Epub 2008 Jul 17. EMBO J. 2008. PMID: 18636090 Free PMC article.
References
-
- Ariumi, Y., A. Kaida, M. Hatanaka, and K. Shimotohno. 2001. Functional cross-talk of HIV-1 Tat with p53 through its C-terminal domain. Biochem. Biophys. Res. Commun. 287:556-561. - PubMed
-
- Ariumi, Y., T. Ego, A. Kaida, M. Matsumoto, P. P. Pandolfi, and K. Shimotohno. 2003. Distinct nuclear body components, PML and SMRT, regulate the trans-acting function of HTLV-1 Tax oncoprotein. Oncogene 22:1611-1619. - PubMed
-
- Bridge, A. J., S. Pebernard, A. Ducraux, A.-L. Nicoulaz, and R. Iggo. 2003. Induction of an interferon response by RNAi vectors in mammalian cells. Nat. Genet. 34:263-264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
