

Review
doi: 10.1016/j.cell.2007.07.039.
Antiviral immunity directed by small RNAs
Affiliations
- PMID: 17693253
- PMCID: PMC2703654
- DOI: 10.1016/j.cell.2007.07.039
Item in Clipboard
Review
Antiviral immunity directed by small RNAs
Cell.
.
Abstract
Plants and invertebrates can protect themselves from viral infection through RNA silencing. This antiviral immunity involves production of virus-derived small interfering RNAs (viRNAs) and results in specific silencing of viruses by viRNA-guided effector complexes. The proteins required for viRNA production as well as several key downstream components of the antiviral immunity pathway have been identified in plants, flies, and worms. Meanwhile, viral mechanisms to suppress this small RNA-directed immunity by viruses are being elucidated, thereby illuminating an ongoing molecular arms race that likely impacts the evolution of both viral and host genomes.
Figures
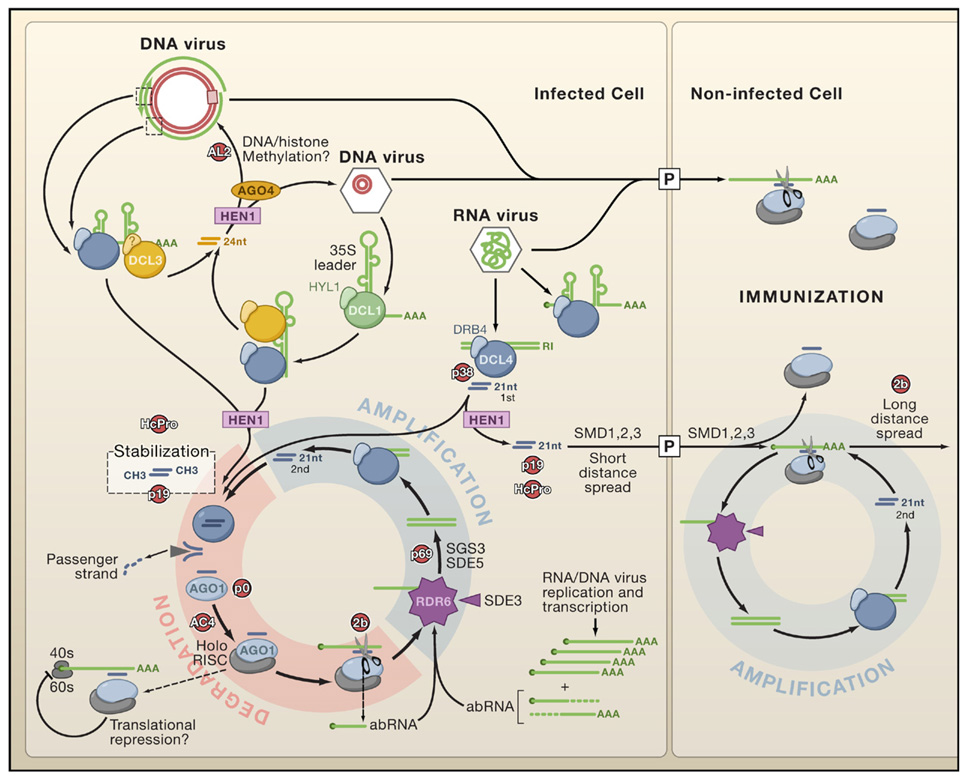
Dicer-like proteins (DCLs) are represented in association with known and unknown cognate double-stranded (ds)RNA-binding proteins (DRBs). Note the indirect implication of DCL1 in viRNA biogenesis from DNA viruses (e.g., Cauliflower mosaic virus) and the putative contribution of DCL3-dependent viRNAs to viral DNA/histone methylation. DCL4 is the primary Dicer to detect RNA viruses and is replaced by DCL2 if suppressed (for example by the VSR P38; see also Figure 3B). AGO1 is presented as a major antiviral slicer, but other AGO paralogs are likely to be involved, potentially also mediating translational repression. All viRNAs are stabilized through HEN1-dependent 2′-O-methylation. The figure shows how primary viRNAs (1st) are amplified into secondary viRNAs (2nd) in the RDR6-dependent pathway. A similar scheme is anticipated with the salicylic acid-induced RDR1 (not shown; Diaz-Pendon et al., 2007). Aberrant (ab) viral mRNAs lacking a cap or polyA tail (AAA) can enter RNA-dependent RNA polymerase pathways independently of 1st viRNA synthesis. A DCL4-dependent silencing signal (arbitrarily depicted as free 21 nucleotide viRNAs) moves through the plasmodesmata (P) to immunize neighboring cells. Movement may be enhanced through further rounds of amplification involving viral transcripts that enter immunized cells. VSRs and potential endogenous silencing suppressors (red) represent genetic rather than direct physical interactions with host silencing components.
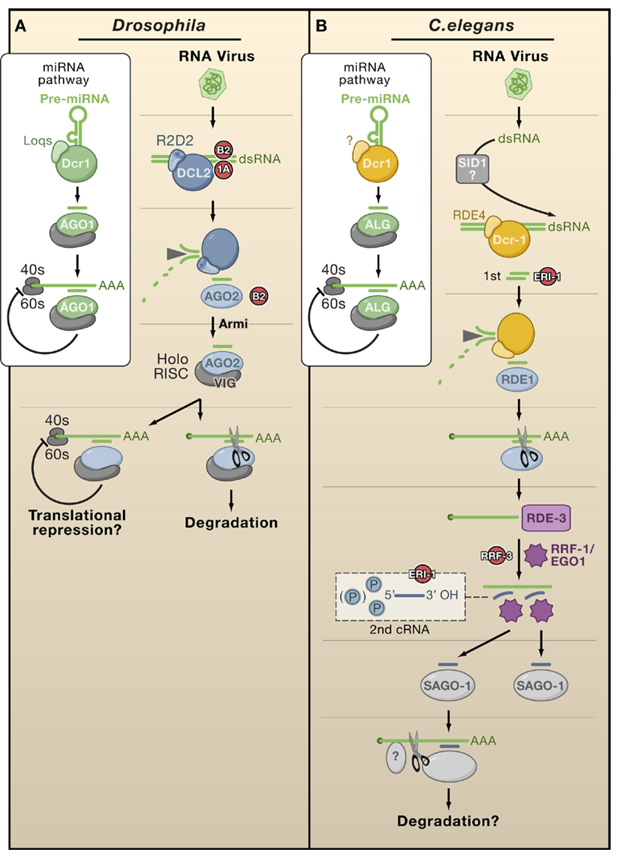
(A) The Drosophila pathway is conceptually similar to a linear antiviral silencing pathway in plants. Although R2D2 heterodimerizes with Dcr2, it is required for loading but not dicing of viRNA; the Armitage (Armi) protein allows assembly of the RISC holoenzyme. The box illustrates the involvement of Dcr1 and AGO1 in the miRNA pathway leading to translational repression. This pathway can be disrupted at multiple points by VSRs (red). (B) Antiviral silencing in C. elegans has been inferred through studies of artificial infection systems. ALG, RDE-1, and SAGO are worm AGOs that recruit miRNAs, 1st siRNAs, and 2nd siRNAs, respectively. RRF-1 is thought to produce 2nd siRNAs or to copy RNAs (cRNAs) directly from RDE3-stabilized templates. SID-1 may possibly take up viral dsRNAs into cells. These pathways can be disrupted at multiple points by endogenous suppressors (red).
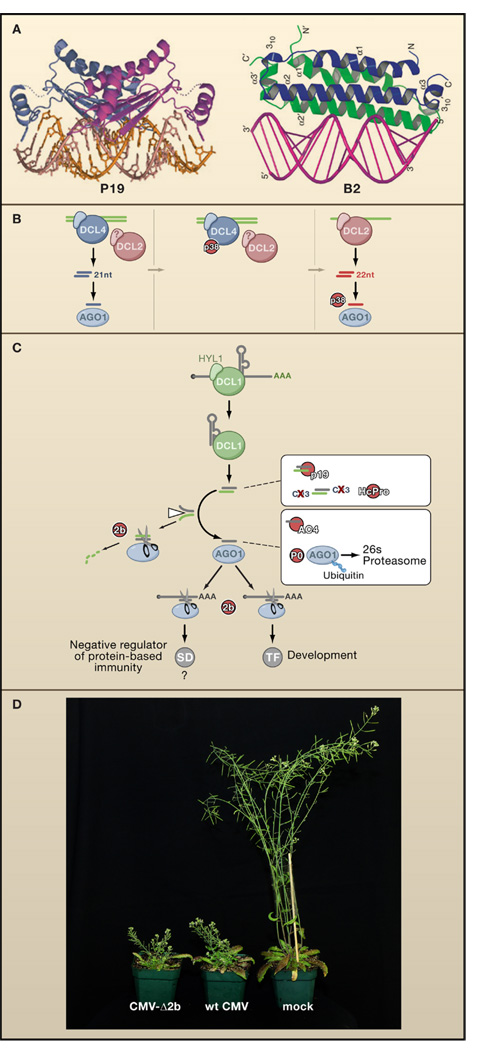
(A) The VSRs P19, encoded by Cymbidium ringspot virus, and B2, encoded by flock-house virus, both bind dsRNA but with very different structural requirements. P19 acts as a head-to-tail homodimer that binds to and specifically measures 21 bp duplexes that are the products of DCL4. In contrast, B2 forms a four-helix bundle that binds to one face of an A-form RNA duplex, independent of its length. P19 in complex with siRNA, reprinted by permission from MacMillan Publishers Ltd: Nature (98), copyright 2003, and B2 in complex with dsRNA, adapted by permission from MacMillan Publisher Ltd: Nat. Struct. Mol. Biol. (109), copyright 2005. (B) Inhibition of DCL4 by the P38 VSR of Turnip crinkle virus reveals the redundant antiviral function of DCL2 which generates 22 nucleotide instead of 21 nucleotide viRNAs in Turnip crinkle virus-infected Arabidopsis. The antiviral activity of DCL2-dependent viRNAs is in turn further compromised by P38, possibly through inhibition of AGO1. (C) Transgenically expressed VSRs interfere with the Arabidopsis DCL1-dependent miRNA pathway. In contrast to the genetic interactions in Figure 1, these interactions are likely to be physical. VSRs may interfere both with developmental programs mediated by transcription factors (TF) and innate immune pathways negatively regulated by suppressors of defense (SD). (D) Both wild-type Cucumber mosaic virus (middle) and a version lacking the VSR 2b (CMV-Δ2b, left) induce similarly severe stunting symptoms in dcl2/dcl4 double mutant plants, demonstrating that the VSR is dispensable for infection and disease induction in a host defective in small RNA-directed immunity (Diaz-Pendon et al., 2007). Image courtesy of R. Lu.
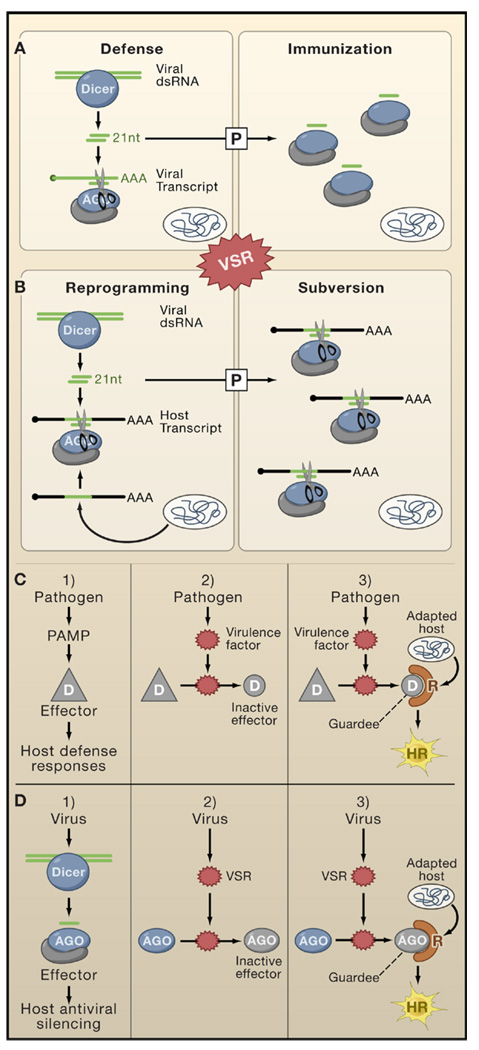
Complex and multilayered interactions exist between RNA silencing pathways, VSRs, and other plant immune pathways. (A and B) This model depicts the possibility that viRNAs produced in virally-infected plants not only contribute to antiviral defense (upper panels) but might sometimes benefit viruses if viRNAs share sequence homologies with host transcripts (lower panels). The model also depicts the possibility that immunization and subversion by viRNAs might operate in cells ahead of the infection front. The extent of defense and subversion is influenced by the timing and level of VSR expression. (C) This model illustrates the guard hypothesis, which proposes that suppression of PAMP-elicited basal defense responses in plants by pathogens’ effector proteins is detected by dedicated host-encoded R proteins, sometimes resulting in a hypersensitive response (HR). (D) An adaptation of the guard hypothesis model explains how VSRs might elicit a hypersensitive response in specific plant ecotypes. The model entails that some effectors of antiviral silencing (e.g., AGO1) are modified by VSRs (such as P0 of poleroviruses; Figure 3C).
Similar articles
-
Antiviral innate immune response of RNA interference.J Infect Dev Ctries. 2014 Jul 14;8(7):804-10. doi: 10.3855/jidc.4187. J Infect Dev Ctries. 2014. PMID: 25022288 Review.
-
Virus counterdefense: diverse strategies for evading the RNA-silencing immunity.Annu Rev Microbiol. 2006;60:503-31. doi: 10.1146/annurev.micro.60.080805.142205. Annu Rev Microbiol. 2006. PMID: 16768647 Free PMC article. Review.
-
Induction and suppression of RNA silencing: insights from viral infections.Nat Rev Genet. 2005 Mar;6(3):206-20. doi: 10.1038/nrg1555. Nat Rev Genet. 2005. PMID: 15703763 Review.
-
RNA-based antiviral immunity.Nat Rev Immunol. 2010 Sep;10(9):632-44. doi: 10.1038/nri2824. Epub 2010 Aug 13. Nat Rev Immunol. 2010. PMID: 20706278 Review.
-
Plant RNA silencing in viral defence.Adv Exp Med Biol. 2011;722:39-58. doi: 10.1007/978-1-4614-0332-6_3. Adv Exp Med Biol. 2011. PMID: 21915781 Review.
Cited by
-
Characterization by Small RNA Sequencing of Taro Bacilliform CH Virus (TaBCHV), a Novel Badnavirus.PLoS One. 2015 Jul 24;10(7):e0134147. doi: 10.1371/journal.pone.0134147. eCollection 2015. PLoS One. 2015. PMID: 26207896 Free PMC article.
-
Exogenous application of double-stranded RNA molecules from TMV p126 and CP genes confers resistance against TMV in tobacco.Planta. 2016 Oct;244(4):961-9. doi: 10.1007/s00425-016-2567-6. Epub 2016 Jul 25. Planta. 2016. PMID: 27456838
-
WRKY8 transcription factor functions in the TMV-cg defense response by mediating both abscisic acid and ethylene signaling in Arabidopsis.Proc Natl Acad Sci U S A. 2013 May 21;110(21):E1963-71. doi: 10.1073/pnas.1221347110. Epub 2013 May 6. Proc Natl Acad Sci U S A. 2013. PMID: 23650359 Free PMC article.
-
Comparative analysis among the small RNA populations of source, sink and conductive tissues in two different plant-virus pathosystems.BMC Genomics. 2015 Feb 22;16(1):117. doi: 10.1186/s12864-015-1327-5. BMC Genomics. 2015. PMID: 25765188 Free PMC article.
-
Dynamics of defense-related components in two contrasting genotypes of tomato upon infection with Tomato Leaf Curl New Delhi Virus.Mol Biotechnol. 2012 Oct;52(2):140-50. doi: 10.1007/s12033-011-9481-8. Mol Biotechnol. 2012. PMID: 22161255
References
-
- Anandalakshmi R, Marathe R, Ge X, Herr JM, Mau C, Mallory A, Pruss G, Bowman L, Vance VB. A calmodulin-related protein that suppresses posttranscriptional gene silencing in plants. Science. 2000;290:142–144. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Bian XY, Rasheed MS, Seemanpillai MJ, Rezaian MA. Analysis of Silencing escape of Tomato leaf curl virus: an evaluation of the role of DNA methylation. Mol. Plant Microbe Interact. 2006;19:614–624. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
