

Sorting of Drosophila small silencing RNAs
- PMID: 17662944
- PMCID: PMC2841505
- DOI: 10.1016/j.cell.2007.05.057
Sorting of Drosophila small silencing RNAs
Abstract
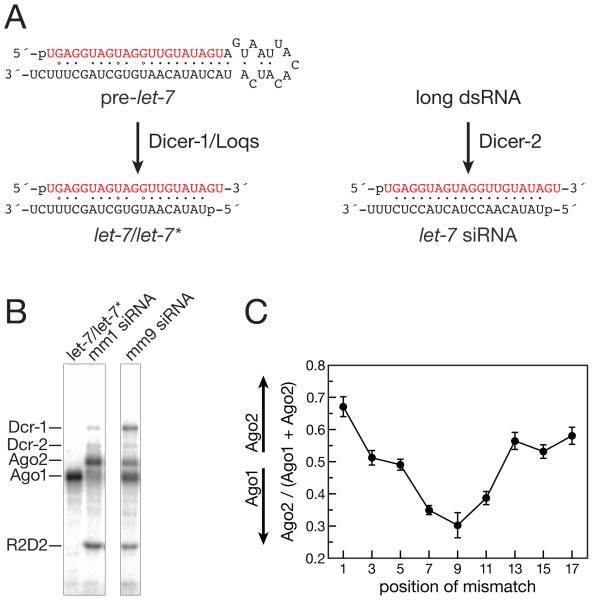
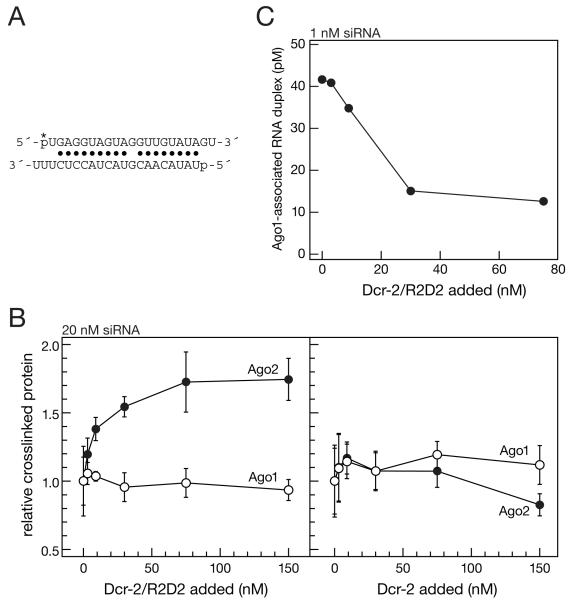
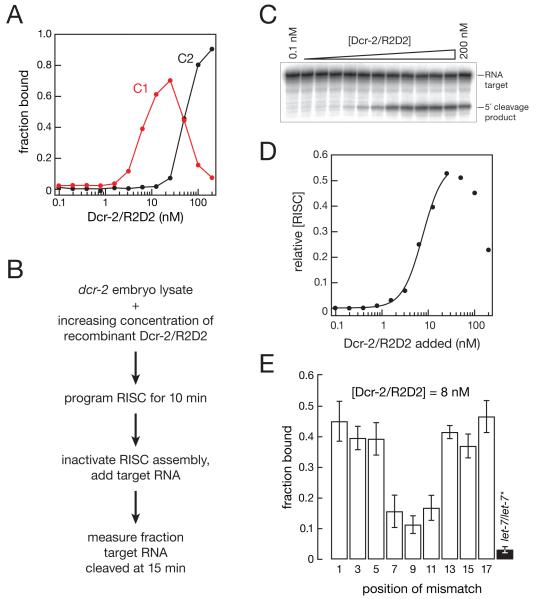
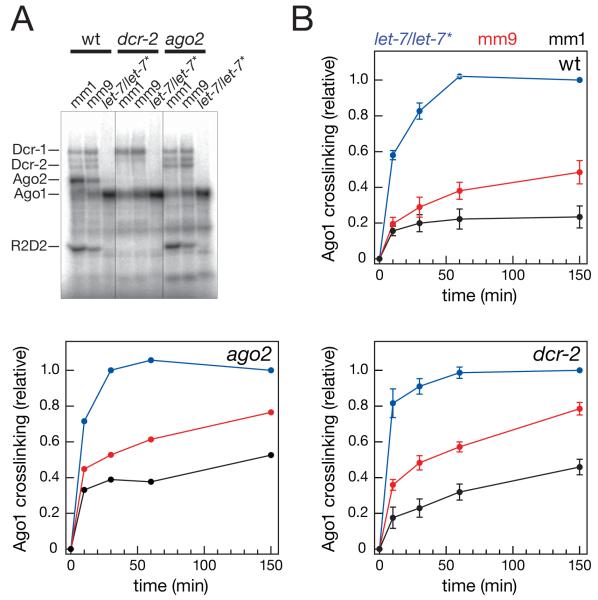
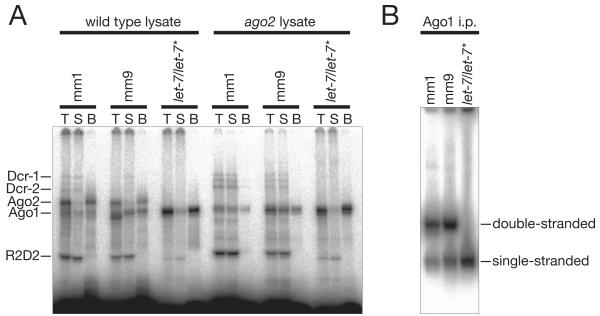
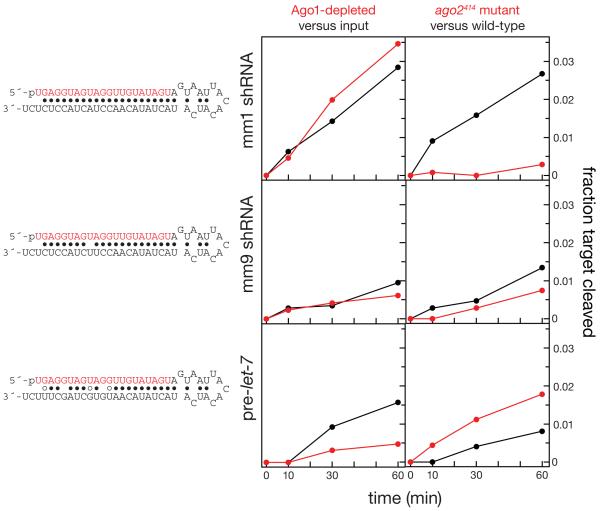
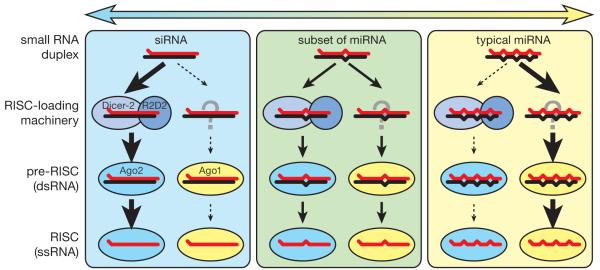
In Drosophila, small interfering RNAs (siRNAs), which direct RNA interference through the Argonaute protein Ago2, are produced by a biogenesis pathway distinct from microRNAs (miRNAs), which regulate endogenous mRNA expression as guides for Ago1. Here, we report that siRNAs and miRNAs are sorted into Ago1 and Ago2 by pathways independent from the processes that produce these two classes of small RNAs. Such small-RNA sorting reflects the structure of the double-stranded assembly intermediates--the miRNA/miRNA( *) and siRNA duplexes--from which Argonaute proteins are loaded. We find that the Dcr-2/R2D2 heterodimer acts as a gatekeeper for the assembly of Ago2 complexes, promoting the incorporation of siRNAs and disfavoring miRNAs as loading substrates for Drosophila Ago2. A separate mechanism acts in parallel to favor miRNA/miRNA( *) duplexes and exclude siRNAs from assembly into Ago1 complexes. Thus, in flies small-RNA duplexes are actively sorted into Argonaute-containing complexes according to their intrinsic structures.
Figures
Similar articles
-
Sorting of Drosophila small silencing RNAs partitions microRNA* strands into the RNA interference pathway.RNA. 2010 Jan;16(1):43-56. doi: 10.1261/rna.1972910. Epub 2009 Nov 16. RNA. 2010. PMID: 19917635 Free PMC article.
-
R2D2 organizes small regulatory RNA pathways in Drosophila.Mol Cell Biol. 2011 Feb;31(4):884-96. doi: 10.1128/MCB.01141-10. Epub 2010 Dec 6. Mol Cell Biol. 2011. PMID: 21135122 Free PMC article.
-
Drosophila microRNAs are sorted into functionally distinct argonaute complexes after production by dicer-1.Cell. 2007 Jul 27;130(2):287-97. doi: 10.1016/j.cell.2007.05.056. Cell. 2007. PMID: 17662943 Free PMC article.
-
Small RNA sorting: matchmaking for Argonautes.Nat Rev Genet. 2011 Jan;12(1):19-31. doi: 10.1038/nrg2916. Epub 2010 Nov 30. Nat Rev Genet. 2011. PMID: 21116305 Free PMC article. Review.
-
MicroRNAs: Loquacious speaks out.Curr Biol. 2005 Aug 9;15(15):R603-5. doi: 10.1016/j.cub.2005.07.044. Curr Biol. 2005. PMID: 16085484 Review.
Cited by
-
Functional parameters of Dicer-independent microRNA biogenesis.RNA. 2012 May;18(5):945-57. doi: 10.1261/rna.032938.112. Epub 2012 Mar 29. RNA. 2012. PMID: 22461413 Free PMC article.
-
Distinct mechanisms for microRNA strand selection by Drosophila Argonautes.Mol Cell. 2009 Nov 13;36(3):431-44. doi: 10.1016/j.molcel.2009.09.027. Mol Cell. 2009. PMID: 19917251 Free PMC article.
-
Rice Dwarf Virus Small RNA Profiles in Rice and Leafhopper Reveal Distinct Patterns in Cross-Kingdom Hosts.Viruses. 2019 Sep 12;11(9):847. doi: 10.3390/v11090847. Viruses. 2019. PMID: 31547224 Free PMC article.
-
Search for limiting factors in the RNAi pathway in silkmoth tissues and the Bm5 cell line: the RNA-binding proteins R2D2 and Translin.PLoS One. 2011;6(5):e20250. doi: 10.1371/journal.pone.0020250. Epub 2011 May 26. PLoS One. 2011. PMID: 21637842 Free PMC article.
-
Mosquito RNAi is the major innate immune pathway controlling arbovirus infection and transmission.Future Microbiol. 2011 Mar;6(3):265-77. doi: 10.2217/fmb.11.11. Future Microbiol. 2011. PMID: 21449839 Free PMC article. Review.
References
-
- Baulcombe D. RNA silencing in plants. Nature. 2004;431:356–363. - PubMed
-
- Bentwich I, Avniel A, Karov Y, Aharonov R, Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, Sharon E, Spector Y, Bentwich Z. Identification of hundreds of conserved and nonconserved human microRNAs. Nat. Genet. 2005;37:766–770. - PubMed
-
- Berezikov E, Thuemmler F, van Laake LW, Kondova I, Bontrop R, Cuppen E, Plasterk RH. Diversity of microRNAs in human and chimpanzee brain. Nat. Genet. 2006;38:1375–1377. - PubMed
-
- Berezikov E, Guryev V, van de Belt J, Wienholds E, Plasterk RH, Cuppen E. Phylogenetic shadowing and computational identification of human microRNA genes. Cell. 2005;120:21–24. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
