

Characterization of EBV gB indicates properties of both class I and class II viral fusion proteins
- PMID: 17655906
- PMCID: PMC2131761
- DOI: 10.1016/j.virol.2007.06.031
Characterization of EBV gB indicates properties of both class I and class II viral fusion proteins
Abstract
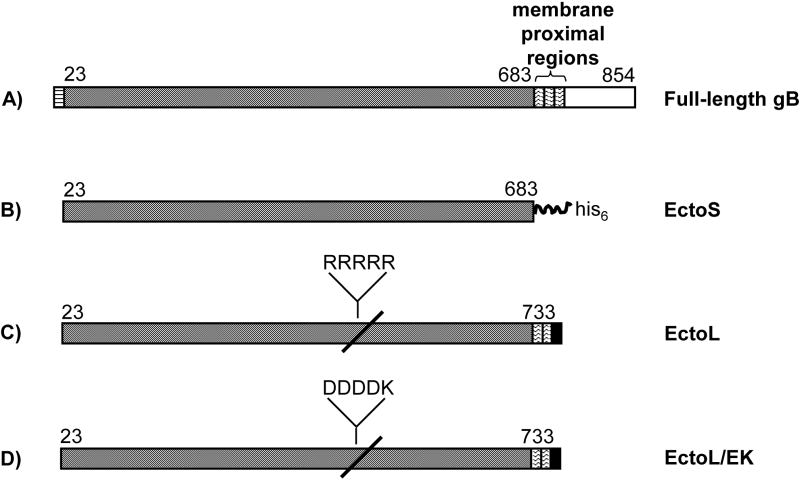
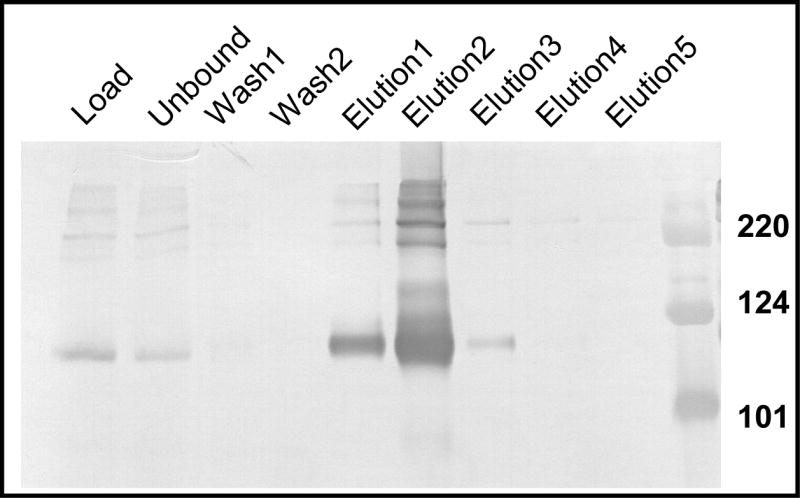
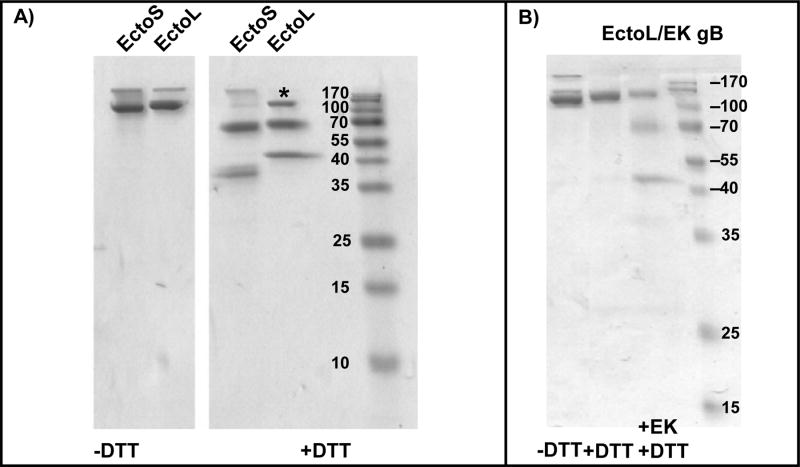
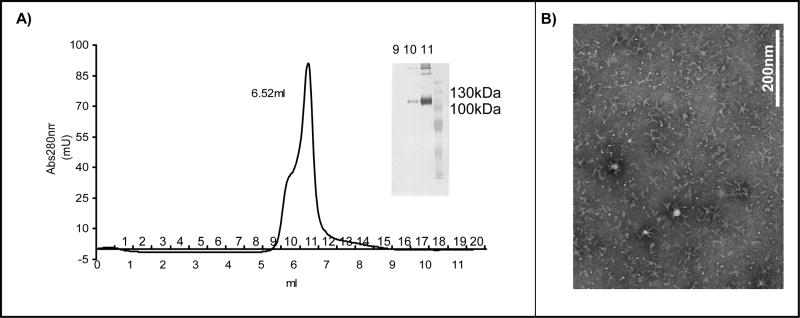
To gain insight into Epstein-Barr virus (EBV) glycoprotein B (gB), recombinant, secreted variants were generated. The role of putative transmembrane regions, the proteolytic processing and the oligomerization state of the gB variants were investigated. Constructs containing 2 of 3 C-terminal hydrophobic regions were secreted, indicating that these do not act as transmembrane anchors. The efficiency of cleavage of the gB furin site was found to depend on the nature of C-terminus. All of the gB constructs formed rosette structures reminiscent of the postfusion aggregates formed by other viral fusion proteins. However, substitution of putative fusion loop residues, WY(112-113) and WLIY(193-196), with less hydrophobic amino acids from HSV-1 gB, produced trimeric protein and abrogated the ability of the EBV gB ectodomains to form rosettes. These data demonstrate biochemical features of EBV gB that are characteristic of other class I and class II viral fusion proteins, but not of HSV-1 gB.
Figures

Similar articles
-
Hydrophobic residues that form putative fusion loops of Epstein-Barr virus glycoprotein B are critical for fusion activity.J Virol. 2007 Sep;81(17):9596-600. doi: 10.1128/JVI.00758-07. Epub 2007 Jun 6. J Virol. 2007. PMID: 17553877 Free PMC article.
-
Modulation of Epstein-Barr virus glycoprotein B (gB) fusion activity by the gB cytoplasmic tail domain.mBio. 2013 Jan 22;4(1):e00571-12. doi: 10.1128/mBio.00571-12. mBio. 2013. PMID: 23341550 Free PMC article.
-
Analysis of Epstein-Barr virus glycoprotein B functional domains via linker insertion mutagenesis.J Virol. 2009 Jan;83(2):734-47. doi: 10.1128/JVI.01817-08. Epub 2008 Nov 5. J Virol. 2009. PMID: 18987135 Free PMC article.
-
Structural and Mechanistic Insights into the Tropism of Epstein-Barr Virus.Mol Cells. 2016 Apr 30;39(4):286-91. doi: 10.14348/molcells.2016.0066. Epub 2016 Apr 6. Mol Cells. 2016. PMID: 27094060 Free PMC article. Review.
-
[The entry of Epstein-Barr virus into B lymphocytes and epithelial cells during infection].Bing Du Xue Bao. 2014 Jul;30(4):476-82. Bing Du Xue Bao. 2014. PMID: 25272606 Review. Chinese.
Cited by
-
Structure-based design of a soluble human cytomegalovirus glycoprotein B antigen stabilized in a prefusion-like conformation.Proc Natl Acad Sci U S A. 2024 Sep 10;121(37):e2404250121. doi: 10.1073/pnas.2404250121. Epub 2024 Sep 4. Proc Natl Acad Sci U S A. 2024. PMID: 39231203 Free PMC article.
-
Class III viral membrane fusion proteins.Curr Opin Struct Biol. 2009 Apr;19(2):189-96. doi: 10.1016/j.sbi.2009.02.012. Epub 2009 Apr 6. Curr Opin Struct Biol. 2009. PMID: 19356922 Free PMC article. Review.
-
Raman Multi-Omic Snapshot and Statistical Validation of Structural Differences between Herpes Simplex Type I and Epstein-Barr Viruses.Int J Mol Sci. 2023 Oct 25;24(21):15567. doi: 10.3390/ijms242115567. Int J Mol Sci. 2023. PMID: 37958551 Free PMC article.
-
Epstein-Barr virus glycoprotein gB and gHgL can mediate fusion and entry in trans, and heat can act as a partial surrogate for gHgL and trigger a conformational change in gB.J Virol. 2014 Nov;88(21):12193-201. doi: 10.1128/JVI.01597-14. Epub 2014 Aug 20. J Virol. 2014. PMID: 25142593 Free PMC article.
-
Mutagenesis of varicella-zoster virus glycoprotein B: putative fusion loop residues are essential for viral replication, and the furin cleavage motif contributes to pathogenesis in skin tissue in vivo.J Virol. 2009 Aug;83(15):7495-506. doi: 10.1128/JVI.00400-09. Epub 2009 May 27. J Virol. 2009. PMID: 19474103 Free PMC article.
References
-
- Blewett EL, Misra V. Cleavage of the bovine herpesvirus glycoprotein B is not essential for its function. J Gen Virol. 1991;72:2083–90. - PubMed
-
- Borza CM, Hutt-Fletcher LM. Alternate replication in B cells and epithelial cells switches tropism of Epstein-Barr virus. Nat Med. 2002;8:594–9. - PubMed
-
- Browne H, Bruun B, Minson T. Plasma membrane requirements for cell fusion induced by herpes simplex virus type 1 glycoproteins gB, gD, gH and gL. J Gen Virol. 2001;82:1419–22. - PubMed
-
- Burnette WN. “Western blotting”: electrophoretic transfer of proteins from sodium dodecyl sulfate--polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal Biochem. 1981;112:195–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
