

MyD88-dependent expansion of an immature GR-1(+)CD11b(+) population induces T cell suppression and Th2 polarization in sepsis
- PMID: 17548519
- PMCID: PMC2118626
- DOI: 10.1084/jem.20062602
MyD88-dependent expansion of an immature GR-1(+)CD11b(+) population induces T cell suppression and Th2 polarization in sepsis
Abstract
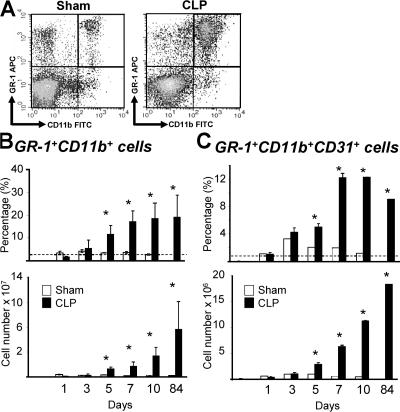
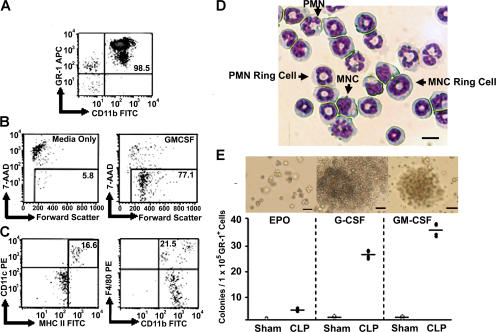
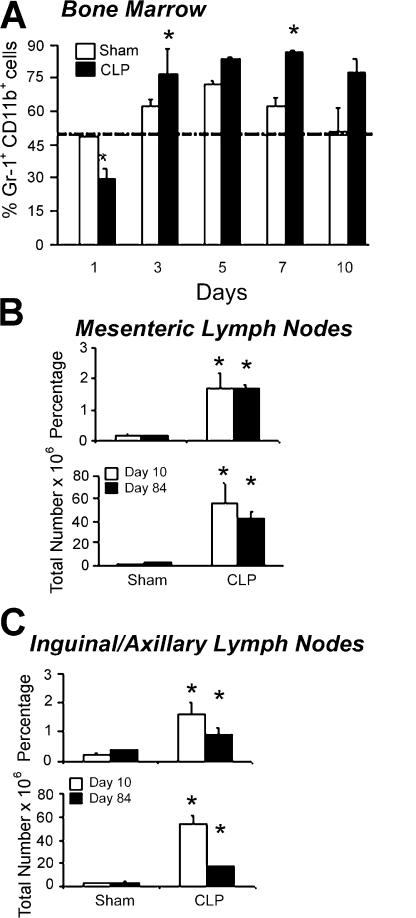
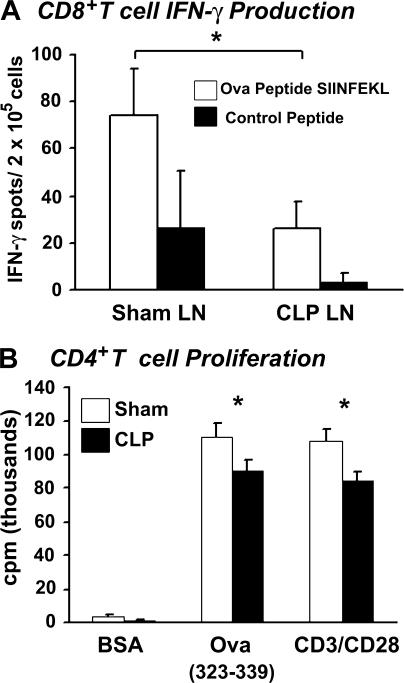
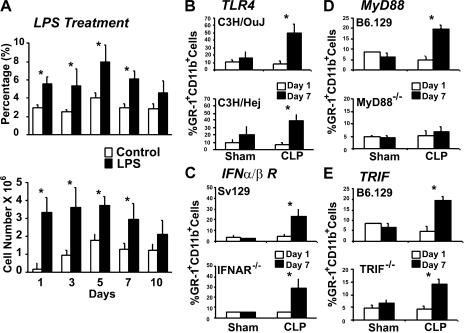
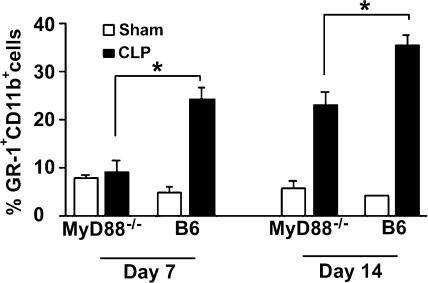
Polymicrobial sepsis alters the adaptive immune response and induces T cell suppression and Th2 immune polarization. We identify a GR-1(+)CD11b(+) population whose numbers dramatically increase and remain elevated in the spleen, lymph nodes, and bone marrow during polymicrobial sepsis. Phenotypically, these cells are heterogeneous, immature, predominantly myeloid progenitors that express interleukin 10 and several other cytokines and chemokines. Splenic GR-1(+) cells effectively suppress antigen-specific CD8(+) T cell interferon (IFN) gamma production but only modestly suppress antigen-specific and nonspecific CD4(+) T cell proliferation. GR-1(+) cell depletion in vivo prevents both the sepsis-induced augmentation of Th2 cell-dependent and depression of Th1 cell-dependent antibody production. Signaling through MyD88, but not Toll-like receptor 4, TIR domain-containing adaptor-inducing IFN-beta, or the IFN-alpha/beta receptor, is required for complete GR-1(+)CD11b(+) expansion. GR-1(+)CD11b(+) cells contribute to sepsis-induced T cell suppression and preferential Th2 polarization.
Figures



Similar articles
-
Hierarchy of immunosuppressive strength among myeloid-derived suppressor cell subsets is determined by GM-CSF.Eur J Immunol. 2010 Jan;40(1):22-35. doi: 10.1002/eji.200939903. Eur J Immunol. 2010. PMID: 19941314
-
Myeloid-derived suppressor cell activation by combined LPS and IFN-gamma treatment impairs DC development.Eur J Immunol. 2009 Oct;39(10):2865-76. doi: 10.1002/eji.200939486. Eur J Immunol. 2009. PMID: 19637228
-
Sepsis leads to a reduced antigen-specific primary antibody response.Eur J Immunol. 2012 Feb;42(2):341-52. doi: 10.1002/eji.201141692. Epub 2011 Dec 20. Eur J Immunol. 2012. PMID: 22105154
-
Subsets, expansion and activation of myeloid-derived suppressor cells.Med Microbiol Immunol. 2010 Aug;199(3):273-81. doi: 10.1007/s00430-010-0151-4. Epub 2010 Apr 8. Med Microbiol Immunol. 2010. PMID: 20376485 Review.
-
Derangement of immune responses by myeloid suppressor cells.Cancer Immunol Immunother. 2004 Feb;53(2):64-72. doi: 10.1007/s00262-003-0443-2. Epub 2003 Oct 30. Cancer Immunol Immunother. 2004. PMID: 14593498 Free PMC article. Review.
Cited by
-
Follicular dendritic cell dysfunction contributes to impaired antigen-specific humoral responses in sepsis-surviving mice.J Clin Invest. 2021 Jun 15;131(12):e146776. doi: 10.1172/JCI146776. J Clin Invest. 2021. PMID: 33956665 Free PMC article.
-
Immune suppression by neutrophils and granulocytic myeloid-derived suppressor cells: similarities and differences.Cell Mol Life Sci. 2013 Oct;70(20):3813-27. doi: 10.1007/s00018-013-1286-4. Epub 2013 Feb 20. Cell Mol Life Sci. 2013. PMID: 23423530 Free PMC article. Review.
-
Malat1 regulates PMN-MDSC expansion and immunosuppression through p-STAT3 ubiquitination in sepsis.Int J Biol Sci. 2024 Feb 11;20(4):1529-1546. doi: 10.7150/ijbs.92267. eCollection 2024. Int J Biol Sci. 2024. PMID: 38385073 Free PMC article.
-
The acute immunological response to blood transfusion is influenced by polymicrobial sepsis.Shock. 2012 Dec;38(6):598-606. doi: 10.1097/SHK.0b013e318273621a. Shock. 2012. PMID: 23143057 Free PMC article.
-
Sepsis: multiple abnormalities, heterogeneous responses, and evolving understanding.Physiol Rev. 2013 Jul;93(3):1247-88. doi: 10.1152/physrev.00037.2012. Physiol Rev. 2013. PMID: 23899564 Free PMC article.
References
-
- Hotchkiss, R.S., and I.E. Karl. 2003. The pathophysiology and treatment of sepsis. N. Engl. J. Med. 348:138–150. - PubMed
-
- Hotchkiss, R.S., K.W. Tinsley, P.E. Swanson, R.E. Schmieg Jr., J.J. Hui, K.C. Chang, D.F. Osborne, B.D. Freeman, J.P. Cobb, T.G. Buchman, and I.E. Karl. 2001. Sepsis-induced apoptosis causes progressive profound depletion of B and CD4+ T lymphocytes in humans. J. Immunol. 166:6952–6963. - PubMed
-
- Oberholzer, C., A. Oberholzer, F.R. Bahjat, R.M. Minter, C.L. Tannahill, A. Abouhamze, D. LaFace, B. Hutchins, M.J. Clare-Salzler, and L.L. Moldawer. 2001. Targeted adenovirus-induced expression of IL-10 decreases thymic apoptosis and improves survival in murine sepsis. Proc. Natl. Acad. Sci. USA. 98:11503–11508. - PMC - PubMed
-
- Murphey, E.D., C.Y. Lin, R.W. McGuire, T. Toliver-Kinsky, D.N. Herndon, and E.R. Sherwood. 2004. Diminished bacterial clearance is associated with decreased IL-12 and interferon-gamma production but a sustained proinflammatory response in a murine model of postseptic immunosuppression. Shock. 21:415–425. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
